the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 26 Nov 2024
| 26 Nov 2024
The Eemian Stage interglacial marine transgression in the south-western Baltic region
Karen Luise Knudsen
Gibbard, P. L. and Knudsen, K. L.: The Eemian Stage interglacial marine transgression in the south-western Baltic region, E&G Quaternary Sci. J., 73, 217–237, https://doi.org/10.5194/egqsj-73-217-2024, 2024.
Through detailed investigations, three sequences of marine and associated deposits in the south-west Baltic area are shown to represent the Eemian interglacial transgression. Pollen analyses and foraminiferal and ostracod analyses are combined to provide the evolution of relative sea-level change. An independent chronology is established by correlation of the pollen record with that from the Bispingen laminated lake deposits in north Germany. Additional published sites from the area are included in a wider reconstruction of events. Ostracod analyses indicate that freshwater lakes initially existed in the basin prior to the marine inundation. This was followed by an early marine transgression. In the south-west Baltic the first marine ingression took place at the base of regional pollen zone E3, with fully marine conditions established in E4 continuing into the lower part of E5. The regression of the sea during zone E6 was followed by freshwater deposition in E7. The linking of the sequences in the south-west Baltic demonstrates that the Eemian transgression occurred both through northern Germany and through the Danish belts–Kattegat seaway, indicating that connections to the North Sea and North Atlantic were present in the area during the maximum interglacial sea-level highstand.
Eine detaillierte Untersuchung von drei marinen Sequenzen und damit verbundenen Ablagerungen im südwestlichen Ostseegebiet stellt nachweislich die interglaziale Transgression des Eem Interglazials dar. Mit hilfe von Pollen- sowie Foraminiferen- und Ostrakodenanalysen wird die Entwicklung der Meeresspiegeländerungen ermittelt. Durch Korrelation der Pollensequenzen mit denen den laminierten Seeablagerungen von Bispingen in Norddeutschland kann eine unabhängige Chronologie erstellt werden. Weitere veröffentlichte regionale Vorkommen werden in eine umfassende Rekonstruktion der eemzeitlichen Abläufe einbezogen. Ostrakodenanalysen deuten darauf hin, dass es vor dem Meeresspiegelanstieg zunächst Süßwassersedimente im Becken gab. Darauf folgte ein frühe Meerestransgression, die in der südwestlichen Ostsee stattfand. Der erste Meereseinbruch ereignete sich zu Beginn der regionalen Pollenzone E3 statt, wobei in E4 vollständig marine Bedingungen herrschten und sich bis in den Beginn von E5 fortsetzten. Dem Rückzug des Meeres in der Zone E6 folgte die Ablagerung von Süßwassersedimenten in E7. Die Verknüpfung der norddeutschen Sequenzen mit weiteren der (süd)westlichen Ostsee, dass die Eem-Transgression sowohl durch das Gebiet des Nord-Ostseekanals als auch durch den dänischen Belt-Kattegat-Seeweg erfolgte, was darauf hindeutet, dass in diesem Gebiet während der Zeit des interglazialen Meeresspiegelmaximums, eine Verbindung zur Nordsee bestand.
- Article
(10756 KB) - Full-text XML
- BibTeX
- EndNote





The Last Interglacial (Late Pleistocene Eemian Stage, ∼ Marine Isotope Substage 5e) has long been recognized as a valuable analogue for understanding the impact of present ongoing and projected climatic warming and questions concerning sea-level change. In particular, the high global sea level during the Eemian Stage (CLIMAP project members, 1984; Shackleton, 1987; Cohen et al., 2022), together with the isostatic depression following from the Late Saalian Stage glaciation, allowed a transgression over large coastal areas along the Baltic Sea coasts, which caused a much more extensive distribution of marine deposits in the Eemian than during the Holocene. Marine Eemian-age deposits therefore occur widely in the western Baltic area. Often these deposits are exposed as glacially dislocated or upthrust sediments in coastal cliffs in the area, such as at classic sites including Ristinge Klint (Kristensen et al., 2000; Knudsen et al., 2011); Gammelmarke, often called Stensigmose (Jessen and Milthers, 1928; Konradi, 1976; Murray and Funder, 2003); and Mommark (Knudsen and Gibbard, 2006) (Figs. 1, 2). Undisturbed marine Eemian-age deposits are found in areas outside the Weichselian Main Stationary Line in south-west Denmark and north-west Germany, for instance at Højer (Penney, 1989) and Dagebüll (Winn et al., 2000). The Eemian deposits in the western Baltic area are generally represented by shallow marine sediments, termed the “Cyprina clay”, formed in inner-shelf, tidal-flat or fjord-like environments.
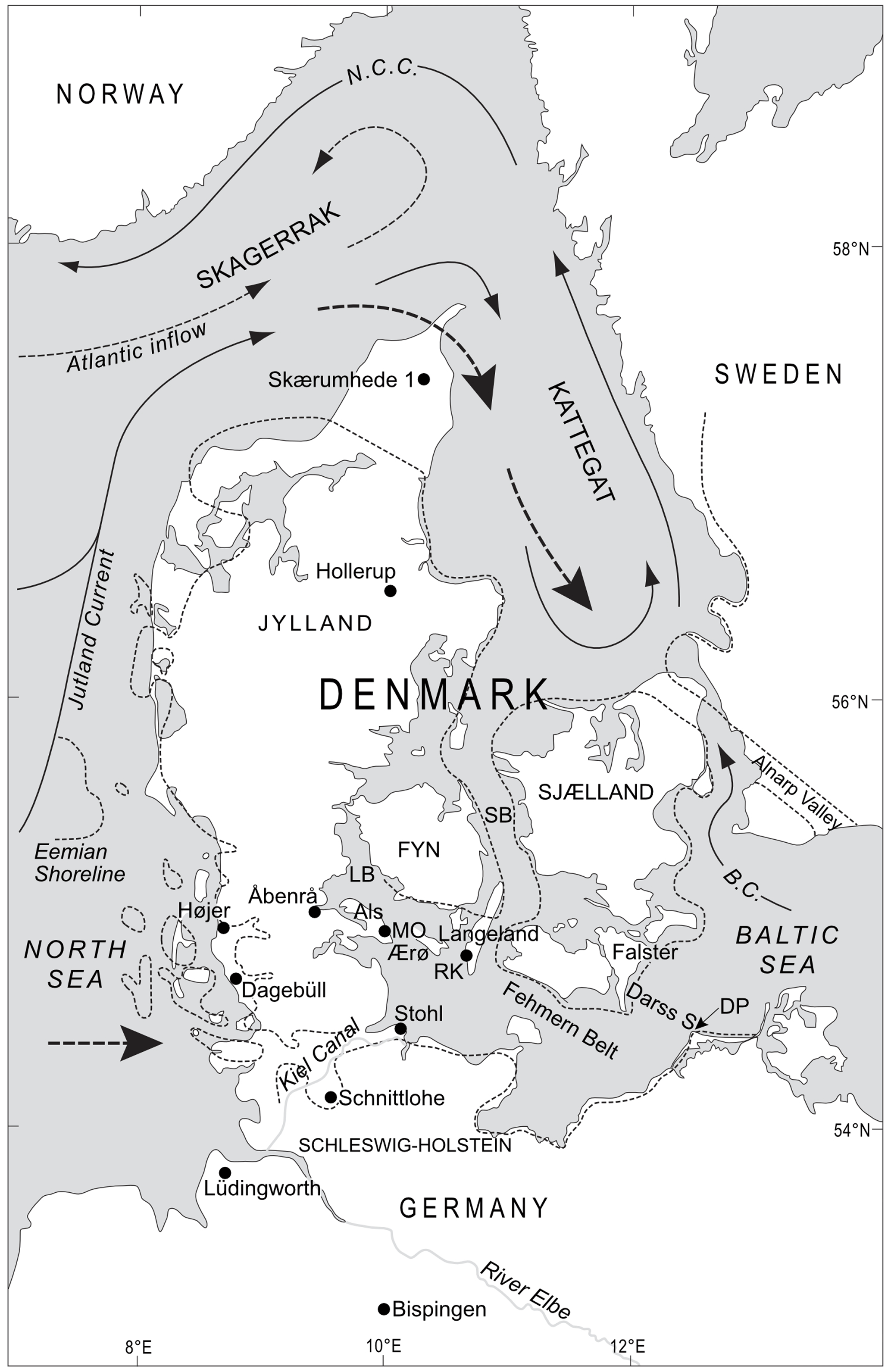
Figure 1Location map of Denmark and northern Germany with an indication (dashed lines) of the suggested maximum Eemian transgression in the area, including the connection across northern Germany around the Kiel Canal, as well as the modern current system. The present-day marine circulation is shown with thin arrows (black arrows are surface currents; stippled arrows are deeper currents). The suggested Eemian inflow of Atlantic water and North Sea water through the Kattegat and Belt seas and through northern Germany, respectively, is indicated by fat stippled lines and large arrows. N.C.C. is Norway Coastal Current; B.C. is Baltic Current; LB is Lillebælt; SB is Storebælt; Darss S. is Darss Sill; DP is Darss Peninsula; RK is Ristinge Klint; Mo is Mommark. (Modified from Kosack and Lange, 1985; Lykke-Andersen and Knudsen, 1991; Seidenkrantz et al., 2000; Funder et al., 2002; and Konradi et al., 2005.)
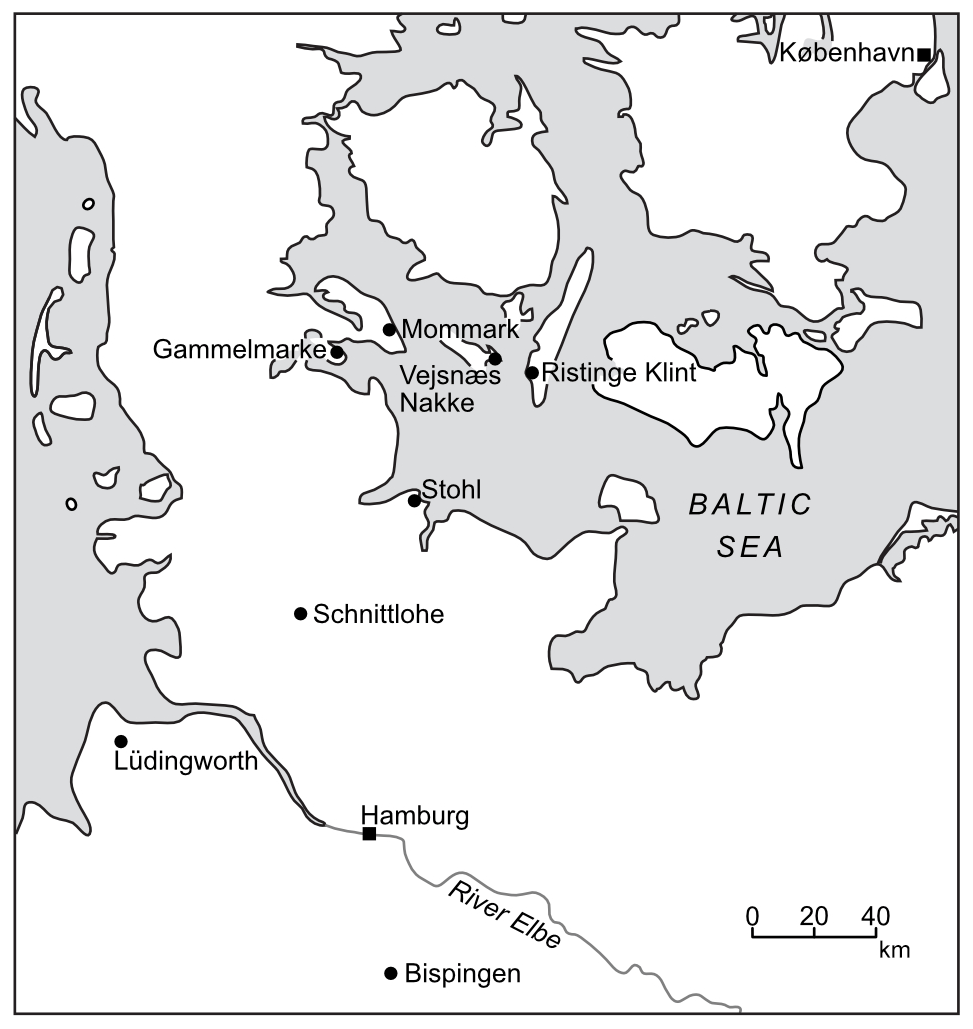
Figure 2Locations of the Eemian Stage sites studied in southern Denmark and northern Germany.
The area close to the Danish belts, with marine connections to the Kattegat, as well as to a potential marine connection across northern Germany through Schleswig-Holstein, makes the western Baltic a key area for the understanding of the palaeoenvironmental changes through the interglacial and in particular the linking of the North Sea and Baltic Sea basins.
The Cyprina clay unit in southern Denmark and northern Germany has been known for more than a century and a half (Forchhammer, 1842), and, by the turn of the century, a correlation with the marine Eemian sediments in the Netherlands was recognized (e.g. Madsen et al., 1908). A large number of scientists have contributed to the discussion of the Cyprina clay both in the Danish and German areas, work that continues to this day (e.g. Jessen and Milthers, 1928; Lafrenz, 1963; Gripp, 1964; Andersen, 1965; Konradi, 1976; Knudsen, 1985; Kubisch and Schönfeld, 1985; Houmark-Nielsen, 1987; Funder et al., 2002; Funder and Balic-Zunic, 2006; Nielsen et al., 2007; Ni et al., 2021).
Further north in Denmark, particularly in northern Jylland, marine Eemian sediments are known from a series of boreholes (120–200 m deep) in a north-west–south-east structurally controlled basin (Lykke-Andersen, 1987). Foraminiferal faunas indicate that the sediments represent continuous deposition from the Late Saalian to Middle Weichselian substages (e.g. Knudsen, 1984, 1992; Knudsen et al., 2009).
The timing of these long-core sequences has been confirmed by both amino-acid geochronology and optically stimulated luminescence (OSL) dating. The former, based on isoleucine epimerization of foraminiferal tests, has clearly shown that the interglacial sediments at Apholm and on Anholt are of Eemian age (Knudsen and Sejrup, 1988), while Miller and Mangerud (1985) reached the same conclusion. OSL dating from a series of deep cores through Eemian-age deposits in northern Jylland was discussed by Larsen et al. (2009). Possible Early Weichselian interstadials have been recognized, overlying the interglacial sediments at Nørre Lyngby, by Glaister and Gibbard (1998) and Kristensen et al. (1998).
The lowermost sediments in the deep boreholes in northern Denmark (e.g. Skærumhede I in Fig. 1 – Knudsen and Lykke-Andersen, 1982; Apholm and Åsted Vest – Knudsen et al., 2009) represent marine conditions that have been referred to the very late Saalian. In the Eemian, Boreal to Boreal–Lusitanian foraminiferal faunas were established in the area at water depths of 60 to over 200 m. At a further 100 km to the south-east, deep-water sediments occur in the same stratigraphical position (e.g. Anholt; Seidenkrantz, 1993a, b). A deep, open embayment therefore was aligned across the northern tip of Jylland into the Kattegat during the Eemian and later. Of particular importance are the Boreal–Lusitanian deep-water foraminiferal species, particularly the Lusitanian Quinqueloculina padana Perconig 1954, which demonstrates that Lusitanian water masses penetrated the whole North Sea basin during this time.
South of the Danish belts many of the small islands are underlain by glacially dislocated Eemian sediments. They mainly consist of marine silty clays (Cyprina clay) and, in the late part of the marine period, sands (the Tapes sand), although freshwater sediments have been identified at several sites predating the transgression event. The molluscan faunas were investigated by Madsen et al. (1908), and the identified faunas contained Tapes senescens and aureus, Mytilus edulis, Ostrea edulis, and Arctica islandica (as Cyprina islandica). Investigations (marine microfossils and pollen) of the classic sequence of ice-pushed sediments at Ristinge Klint by Kristensen et al. (2000) have shown a marine transgression into a freshwater basin at the regional pollen zone E3–E4 transition (Andersen, 1975). The transgression continued and established nearly fully marine conditions in the later part of regional pollen zone E4 (Head, 2007; Knudsen et al., 2011). This record is comparable to that reported from the Kiel Canal area of neighbouring Schleswig-Holstein (Hinsch, 1985; Knudsen, 1985; Kosack and Lange, 1985; Menke, 1985). Whether the transgression took place through the Danish belts or across the southern part of the Danish–German peninsula, or both, was not clear previously, but results from the Kiel Canal area in northern Germany indicate that a connection with the North Sea was present in the area for at least a major part of the interglacial sea-level highstand interval (BALTEEM participants, 2002).
This article presents the results from three sites in the western Baltic area, and, by linking these with the evidence already published from the region, evaluates the nature and relative timing of the marine transgression during the Eemian Stage in the south-western Baltic area and the development of the accompanying environments.
2.1 Pollen and spores
Samples for pollen analysis were prepared using standard chemical methods (West, 1977), modified by using clay dispersion techniques described by Bates et al. (1978). Pollen type nomenclature follows Andrew (1984), together with some types listed in Birks (1973). The pollen counts have been calculated using the pollen sum of total pollen and spores, excluding pre-Pleistocene taxa and dinoflagellate cysts, which were calculated as a percentage of total palynomorphs. This technique has been adopted since the majority of sediments in the sequences analysed are of marine origin, and therefore all the taxa identified have been transported to the depositional site. In particular spores of Sphagnum and ferns have not been excluded since they reflect the local vegetation and can be strongly dominant and overrepresented.
The descriptions of the individual sequences below use the original depths as measured at the time of sampling. In order to avoid any question of confusion over the compatibility of results, wherever possible the same sample interval used for the foraminifera was also used for the palynology. The sequences are subdivided into local pollen assemblage zones (l.p.a.z.'s) to assist with site correlation to regional pollen assemblage zones (r.p.a.z.'s). The sequences are compared and correlated with others in the region and the time span represented determined by comparison to sites of known age from elsewhere in northern Europe.
The palynology of marine sediments allows the sequences to be cross-correlated with those on land from which the development of the interglacial vegetation is well known. Nevertheless, the Eemian (and its equivalents) of the Baltic region is well established from terrestrial sites (Zagwijn, 1983, 1996; Donner, 1995; Aalbersberg and Litt, 1998, among others), and therefore the correlation with the marine sediments is unambiguous.
2.2 Foraminifera and ostracods
Samples for foraminiferal and ostracod analyses were prepared using standard techniques described in Feyling-Hanssen et al. (1971) and Knudsen (1998), dried sediment samples being wet sieved on 0.1 and 1 mm sieves and subsequently concentrated with heavy liquid (C2Cl4). The ostracods were picked from both the heavy and the light 0.1–1 mm fraction. The foraminiferal taxonomy and nomenclature follows Ellis and Messina (1949, with supplements), supplemented by more recent literature from the region. From each sample, a minimum of 300 tests for each group (foraminifera and ostracods) were picked where possible, though in several samples the numbers were too low, and therefore all tests present were counted. Samples with fewer than 50 specimens have been excluded from the percentage diagrams. Here, the combined foraminiferal and ostracod assemblages have been divided into assemblage zones based on visual inspection of faunal changes. Table 1 shows a list of the foraminiferal and ostracod species mentioned in the text with their original references.
Table 1Species list with original references for the foraminiferal and ostracod species mentioned in the text.

2.3 Age model
The timing of the marine deposition throughout the south Baltic region is determined on the basis of pollen stratigraphy. As already noted, each of the records can be closely correlated with pollen sequences from those of the Last Interglacial (Eemian Stage), for example Herning (Jessen and Milthers, 1928), Hollerup (Andersen, 1961, 1975; Björck et al., 2000), Bispingen (Müller, 1974; Lauterbach et al., 2024), Rederstall (Menke and Tynni, 1984), Osterbylund (Thiel et al., 2023) and locations described by Zagwijn (1983, 1996). Indeed, the diagrams can be zoned directly according to the scheme of Jessen and Milthers (1928) and Menke and Tynni (1984) that were developed in Denmark and north-west Germany (Fig. 3). Equally, the successions can be zoned using Andersen's later scheme (1961, 1975) for the Danish lacustrine Eemian Stage parastratotype locality at Hollerup (Fig. 1).
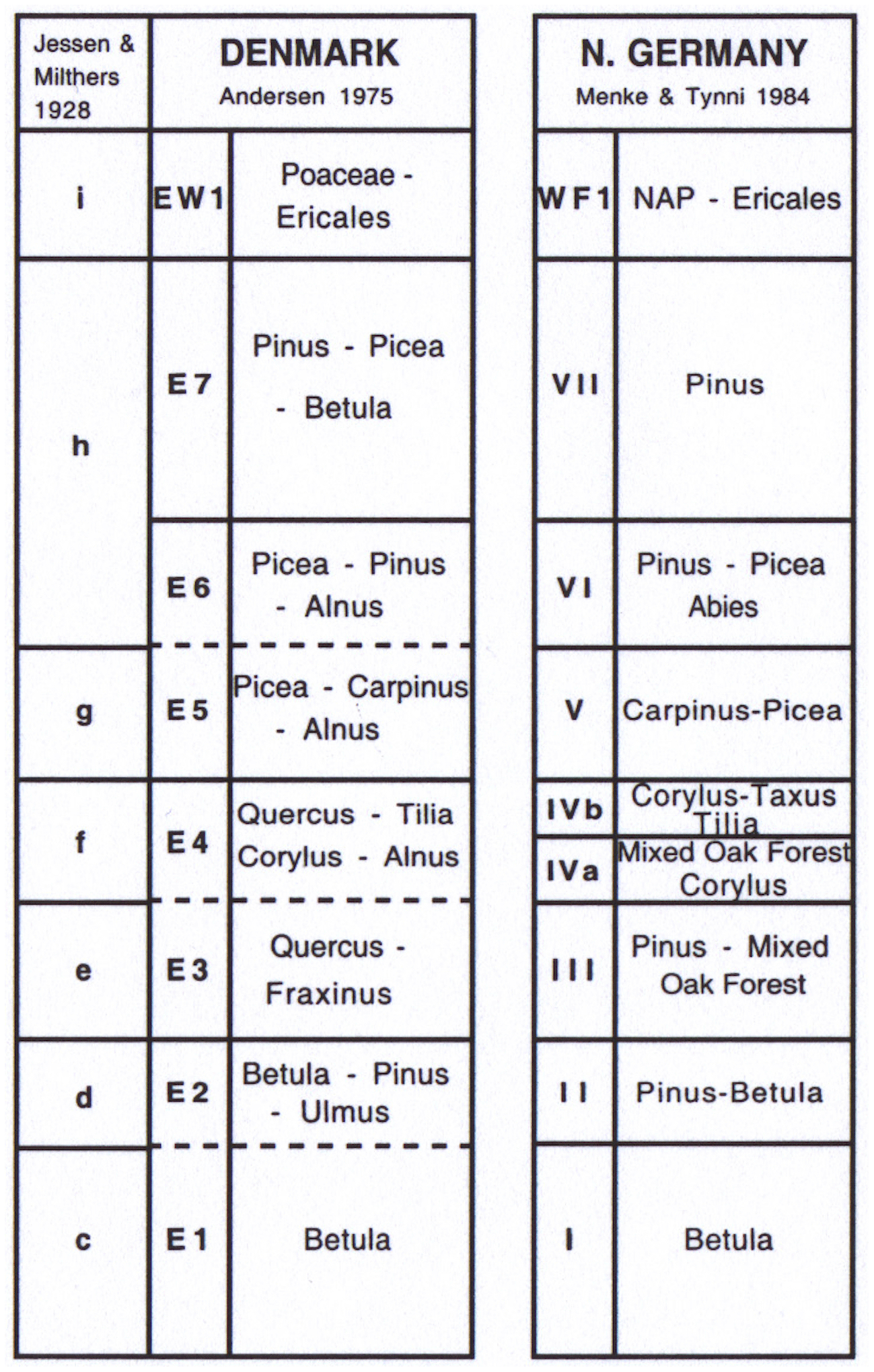
One of the main characteristics of the Eemian Stage in temperate north-west Europe is the apparent uniformity of the vegetational development, shown by the progressive peaks of tree taxa. Consequently, Zagwijn (1996) concluded that the zonal boundaries probably do not differ by more than 200–500 years across the whole of the northern European region. The establishment of the strong correlation of the sites within the south-west Baltic area with the Eemian Stage is important, in particular because this enables the sequences, the majority of which represent fragments of the interglacial interval, to be equated with that at the lacustrine site at Bispingen, Lower Saxony (Figs. 1, 2). Here Müller (1974) and more recently Lauterbach et al. (2024) have described lacustrine sediments that are annually laminated in the first part of the sequence spanning ∼ 3000 years, and the counts have been extrapolated for the rest of the interglacial so that the interglacial appears to span ∼ 15 000 years in total. Therefore, it is possible to use these approximate ages to “date” regional pollen zone boundaries and thus provide a floating chronology for the region (see Zagwijn, 1983, 1996; Larsen et al., 1995; Lauterbach et al., 2024). A palynological correlation of each of the three new sites studied in this region with the zonation of Andersen (1964, 1965) from Hollerup and that of Müller (1974) and Lauterbach et al. (2024) from Bispingen is shown below.
However, one problem is that the Bispingen sequence may not start at the beginning of the interglacial, possibly implying that there may be a few hundred years missing from the age estimates for the stage, hence the need to add or subtract as much as 1000 years from the ages quoted. Nevertheless, remarkably similar estimates have been obtained from other sequences in northern Germany, in particular Quakenbrück (Hahne et al., 1994) and Groß Todtshorn (Caspers, 1997). Turner (2002) and Caspers et al. (2002) further discuss the duration of the Eemian Stage in the region. Given the approximate ages of the pollen zone boundaries, the recently revised reassessment of the duration of the interglacial temperate conditions proposed by Lauterbach et al. (2024) is used as the basis for the chronology.
3.1 The Gammelmarke cliff
With a research record of more than a century (e.g. Gottsche, 1904), the cliffs at Gammelmarke (Figs. 2, 4; often called Stensigmose in earlier literature; 54°52′40.42′′ N, 9°44′27.96′′ E) in southern Jylland are among the classical Eemian sites. The correlation with the Eemian at its type locality in the Netherlands was based on the marine mollusc fauna with a Lusitanic (south-west European) element and the abundance of the extinct bivalve Paphia (formerly Tapes) senescens, which characterizes shallow marine sediments in north-western Europe (Madsen et al., 1908; Nordmann, 1908). This was supported by pollen and plant macrofossil analyses (Jessen and Milthers, 1928) and has been confirmed by more recent work on foraminiferal faunas (Konradi, 1976). The sediments at Gammelmarke have all been disturbed, folded, thrusted and dislocated by the Weichselian ice sheet, which overrode the area and reached its outermost limit ∼ 30 km to the west. However, a general succession of sediments can be identified along the cliff (e.g. Konradi, 1976; Murray and Funder, 2003). At beach level, the sequence begins with Saalian-age till and meltwater sand. The start of the interglacial is seen in the lacustrine Anodonta sand and clay or occasionally peat. After this, the rising sea level caused the sea to inundate the area, reflected in a marine transgression lag, the Mytilus horizon. This is overlain by a ∼ 10 m thick succession of marine sediments coarsening upwards, beginning with offshore mud, the Cyprina clay, named after the bivalve Cyprina (now Arctica) islandica. Gradually this changes to bioturbated shoreface Tapes sand, named after the abundant Tapes (now Paphia) senescens. The Tapes sand is unconformably overlain by a bed of unfossiliferous faintly laminated rust-stained sand, which generally has been interpreted as glaciofluvial meltwater sand from the Weichselian ice sheet. At the top of the succession is a 1–2 m thick till bed.
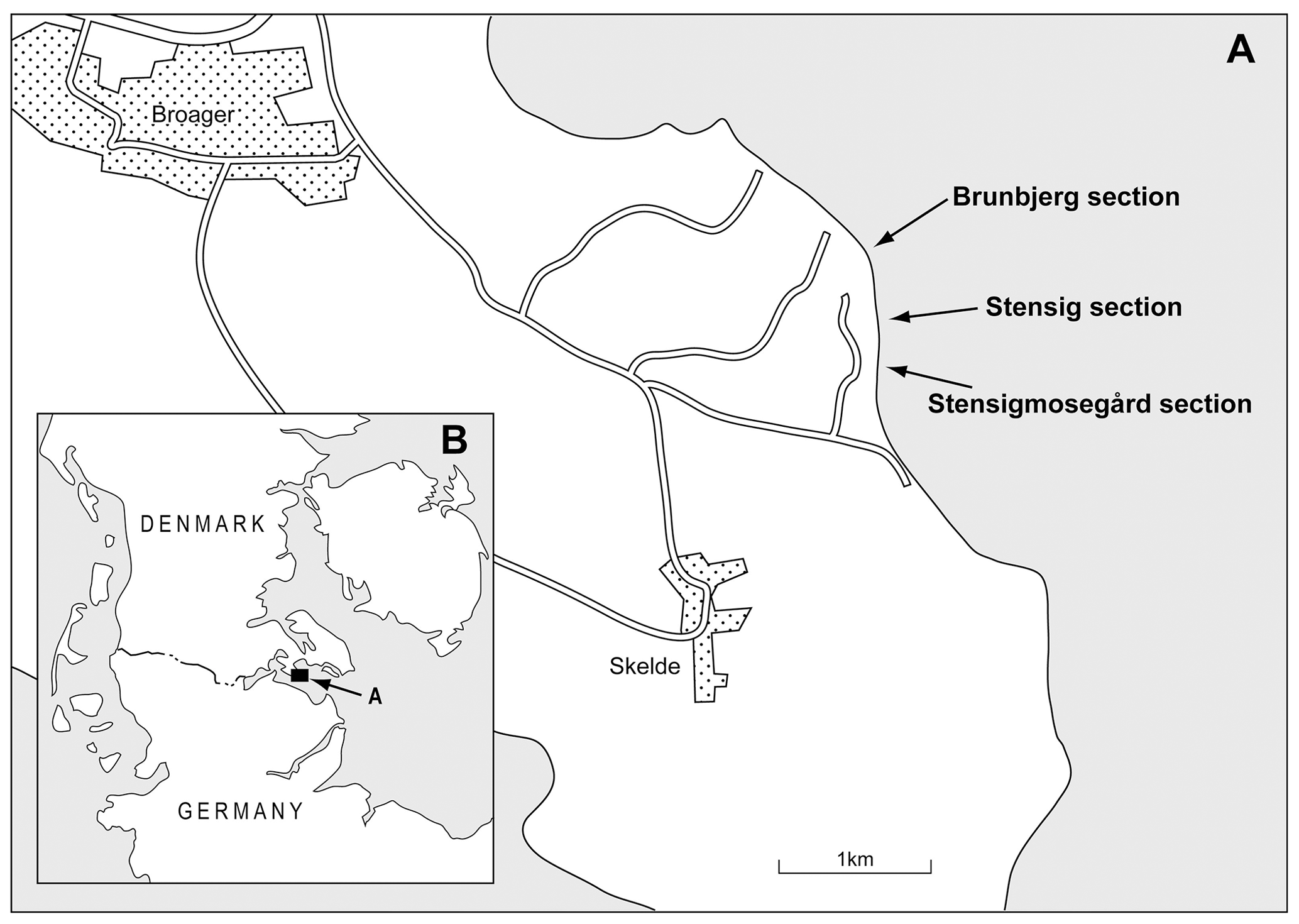
Figure 4Details with locations of three coastal sections at Gammelmarke on the east coast of southern Jylland.
This general sequence can be recognized along the entire cliff, but there is much horizontal variation both in thickness and facies of individual strata. Therefore, the record at Gammelmarke is unusually rich in detail on late Quaternary periglacial and non-glacial environments. Three sections along the cliff have been studied (Fig. 4). In the north, at the Brunbjerg section, closely spaced OSL dating was carried out in order to control the estimate of the duration of marine sedimentation during the Eemian (Murray and Funder, 2003). Some hundred metres to the south, at the Stensig section, a series of samples originally obtained by Ditlefsen (1991) have been analysed for pollen, foraminifera and ostracods in the present study. In the south, the Stensigmosegård section provides new evidence on environmental conditions, particularly for the lacustrine sediments and the time of marine transgression (BALTEEM participants, 2002).
The Brunbjerg section, the northernmost of the exposed sections at Gammelmarke (Fig. 4), was chosen by Murray and Funder (2003) for OSL dating because the sediments at this site have preserved their almost horizontal setting. The age for the sampled sediments from the Late Saalian to the early mid-Eemian was expected to cover the interval ∼ 133 to ∼ 128 ka because (1) pollen analyses at other sections at Gammelmarke (see below) show that non-glaciogenic sedimentation began in the Late Saalian and continued into the early Eemian; (2) the beginning of the Eemian, i.e. the period of forest cover in north-west Europe, is estimated at ∼ 132 ka (Late Saalian and early Eemian sea-level rise; Shackleton, 2000; Funder et al., 2002); and (3) the marine period probably ended when the interglacial sea-level highstand was achieved and isostatic rebound became dominant at about 128 ka (Funder et al., 2002). The results of the dating gave an overall average age of 119 ± 2 ka (n = 22), results that are discussed by Murray and Funder (2003). The Brunbjerg section is located close to Profile II, described by Konradi (1976).
The Stensig section (Fig. 4), which is located close to Profile I of Konradi (1976), was described and dated by Ditlefsen (1991). Here, glaciofluvial silt and sand are overlain by lacustrine sediments, consisting of clay, peat and gyttja, rich in Anodonta shells. This is followed by marine clay with thin sand layers, gradually coarsening upwards to silt and sand. The marine sequence is interrupted by a 40 cm thick cross-bedded sand stratum, rich in gastropod fractions (at 4.35–4.75 m a.s.l., corresponding to 2.25–1.85 m depth in the Stensig section diagrams below). Glacial sediments from the Weichselian cover the marine sequence in the section. The pollen stratigraphy and the foraminiferal and ostracod assemblage zones for the Stensig section are described below.
The Stensigmosegård section in the southern part of the cliff (Fig. 4) deviates from the above-mentioned exposures by the deformations in the Anodonta clay and sand and by the absence of Cyprina clay. After the marine transgression a thin bed of clayey sand is directly overlain by Tapes sand. At this section, Late Saalian till and glaciofluvial deposits are thus overlain by lacustrine and marine deposits representing regional pollen zones E2 and E3 (zone E1 is missing; BALTEEM participants, 2002). The marine transgression occurred in the lowermost part of E3, 300–500 years after the beginning of the Eemian (ca. 131.5 ka).
3.2 Stensig section
3.2.1 Pollen and spores
A total of 11 samples from the Stensig section were analysed for their pollen and spore content. The samples used were also subsampled for foraminiferal and ostracod analysis. The condition of the palynological material was generally poor, with the oxidation of grains a particular problem. The condition of the grains may potentially have biased the pollen assemblages causing those grains which are easily identified, for example Alnus and Pinus, to potentially be overrepresented. This has been considered in the interpretation. The pollen diagram is shown in Fig. 5.
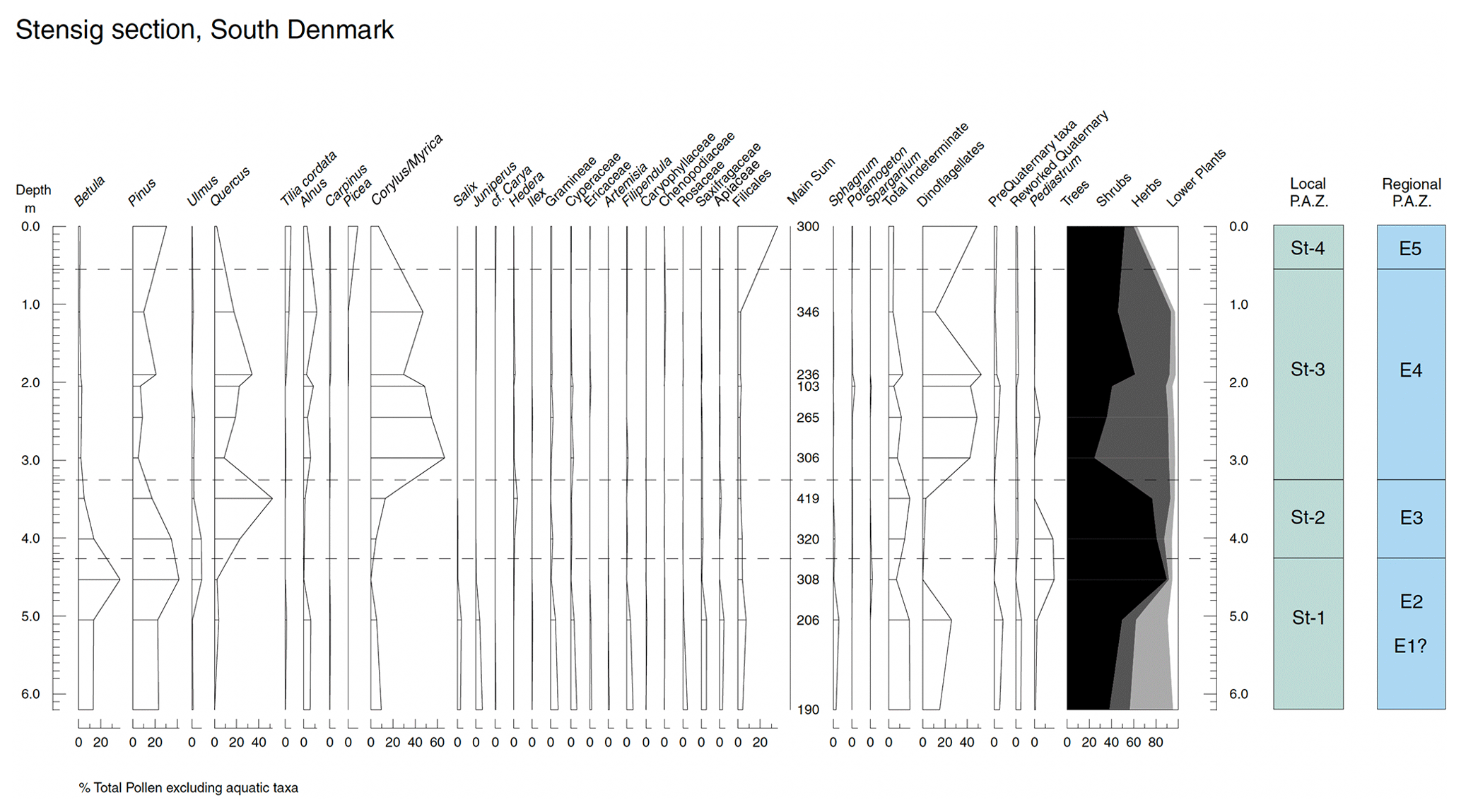
Figure 5Summary pollen diagram from the Stensig section at the Gammelmarke coast cliff, southern Denmark, with correlation to the regional pollen assemblage zones (Andersen, 1961, 1975).
Zone St-1: 620–425 cm, Pinus–Betula l.p.a.z.
This zone is characterized by high proportions of Betula and Pinus (up to 37 % and 41 %, respectively) pollen, with relatively high proportions of herbaceous taxa. The herb pollen includes Poaceae, Cyperaceae, Ericaceae, Filipendula, Rosaceae, Saxifragaceae and Apiaceae, as well as a decrease in percentage occurrence in the uppermost part of the zone. Taken as a whole, the assemblage recorded in this zone reflects an open pine–birch woodland, with surrounding areas of grassland with other herb taxa typical of dry grassy environments, e.g. members of Rosaceae, Saxifragaceae and Apiaceae. Damp soils, probably around watercourses, were populated by Alnus and herbs and mosses which favour wet ground, such as Filipendula and Sphagnum. Reworked pre-Pleistocene taxa are rare, although the association with relatively high (ca. 20 %) proportions of unidentifiable grains, in conjunction with the occurrence of Alnus, may be explained by the inwash of river-borne pollen.
The uppermost level of the zone sees a decrease in importance of herbaceous pollen taxa, together with peaks in Betula and Picea and the apparent immigration of the thermophilous tree taxon Ulmus. This is interpreted to represent a reduction in open grassland and the spread of tree taxa as a response to climatic amelioration.
Zone St-2: 425–325 cm, Quercus l.p.a.z.
The lowermost boundary of this zone is defined on the basis of a sharp increase in Quercus pollen, which reaches a maximum of 52 % in the uppermost part. Although Quercus was consistently present in the previous zone, this was at very low percentages and most likely reflects a background signal through long-distance transport or, more likely, the reworking of earlier interglacial sediments.
The increasing influence of Quercus is reflected in a decline in the frequency of Betula and Picea pollen, which had been the dominant taxa in the previous zone. Ulmus, an important part of the forest vegetation in the early part of the zone, and Corylus pollen percentages show a gradual increase throughout. As with Quercus, grains of Corylus were recorded prior to this increase; in this case, these occurrences probably reflect a combination of reworking and also misidentification of Myrica gale, a plant of moist, boggy sites close to the sites, which has a very similar pollen morphology to that of Corylus. This misidentification could reflect poor pollen preservation in the assemblage.
This zone reflects the development of fully temperate mixed-oak forest, characterized by the dominance of oak, with associated thermophilous tree taxa including Ulmus and an understorey rich in Corylus. The temperate, interglacial character of the vegetation is confirmed further by the occurrence of Hedera and possibly Ilex (represented by a single pollen grain) in this zone, both of which are frost-sensitive. Pine and birch remain important components of the vegetation, although in the case of Pinus it is difficult to define accurately its true extent as a result of potential overrepresentation. However, a further decline in pine frequencies in the subsequent zone suggests that not all the grains recorded here reflect long-distance transport.
Of particular note is the high proportion of Pediastrum recorded in the lowermost part of the zone and the comparative lack of Alnus. Pediastrum is a freshwater alga, common in low-altitude lacustrine environments. In conjunction with the sharp rise in dinoflagellates in the subsequent zone, this suggests that zone St-2 represents a change in the depositional environment and a transition from lacustrine to shallow marine sedimentation, possibly through an estuarine stage.
Zone St-3: 325–55 cm, Corylus l.p.a.z.
While proportions of Corylus pollen showed a slight increase through the previous zone, the first level of St-3 records a rise in Corylus to 66 % of total land pollen and marks the lowermost boundary of the zone. Partly as a result of the closed dataset inherent in pollen percentage diagrams, Quercus frequencies fall sharply to a mean of 20 %. As suggested by the concentration data, this change does not reflect a wholesale replacement of oak forest by Corylus, since hazel would have predominantly grown in the understorey of the mixed-oak forest.
Of the other tree taxa, Ulmus pollen frequencies fall to low levels, while Alnus percentages recover, reflecting growth predominantly along watercourses. River input is also suggested by the occurrence (albeit in very low proportions) of the freshwater taxa Potamogeton and Sparganium. The thermophilous nature of the woodland is emphasized by the immigration of Tilia in the upper part of the zone.
Herb pollen is not abundant and shows little change in type or occurrence from the previous zone. A notable exception is the pollen of Chenopodiaceae, the persistent occurrence of which implies the development of salt-marsh vegetation in a coastal environment through the latter part of the period represented.
Zone St-4: 55–0 cm, Pinus–Picea–Pteropsida l.p.a.z.
This zone is based on only one sample. Although this would not normally be a valid criterion for subdivision, the assemblage is thought to be sufficiently distinct from subsequent samples to merit a tentative separation. The zone is notable for a sharp decline in the occurrence of pollen of thermophilous trees, with Quercus and Corylus in particular being present as less than 10 % of total land pollen.
While Betula pollen shows little change from the previous zone, Pinus increases, and, for the first time in the sequence, Picea pollen occurs at 10 %. Carpinus is also present, albeit at very low proportions, having been recorded in the uppermost part of the previous zone. The assemblage present at this level, in particular the immigration of Picea, suggests the earliest stage of vegetational response to deteriorating soil conditions and the formation of podsols (e.g. Andersen, 1966; Turner and West, 1968; West, 1980). The level on which this zone is defined bears a strong resemblance to the early part of zone Mom-c at Mommark (Kristensen and Knudsen, 2006), and it is reasonable to suppose that, had the sequence continued at the Stensig section, it would have recorded the development of a Carpinus–Picea-dominated woodland, as seen at the former locality.
Herbaceous pollen taxa show little change from the rest of the sequence, but a significant change is recorded in the curve for Pteropsida (Filicales) spores, which increase to 35 % at this level from a previous occurrence of ca. 2 %. Such a dramatic change gives cause to doubt the reliability of an ecological interpretation based on this taxon. There are two possible explanations for the rise: that it reflects a genuine change in the environment or that it reflects the reworking of soils or earlier sediments. The occurrence of Pteropsida (Filicales) spores is difficult to interpret, not least because of their resistance to physical or chemical damage (hence making them a dominant component of reworked assemblages) and the number of habitats which different members of the Pteropsida (Filicales) ferns inhabit. In this case, the possibility that the Pteropsida (Filicales) curve reflects reworking can be possibly discounted because the covariance which would occur with pre-Quaternary taxa under such conditions is not observed. However, the interpretation of the Pteropsida as a genuine member of the vegetational assemblage is supported in this case by the development of a spruce forest. Ferns are essentially plants of damp habitats and are commonly associated with damp shady environments, such as those provided by spruce forest, as opposed to the relatively dry, open oak woodland. Although this interpretation must remain tentative due to the relative lack of information in this zone, it is reasonable to consider that the Pteropsida (Filicales) curve is most probably a genuine reflection of the vegetation at the time of deposition. Nevertheless, another possible interpretation could be that the elevated pine and Pteropsida values are a result of selective pollen preservation leading to the enrichment of these taxa. To a minor extent, this could also apply to spruce given the poor pollen preservation throughout these sediments.
On the basis of the l.p.a.z.'s defined here they are equated to regional pollen assemblage zones (r.p.a.z.'s) as follows: St-1 = E2, St-2 = E3, St-3 = E4 and St-4 = E5 (see Andersen, 1975; Zagwijn, 1983).
3.2.2 Foraminifera and ostracods
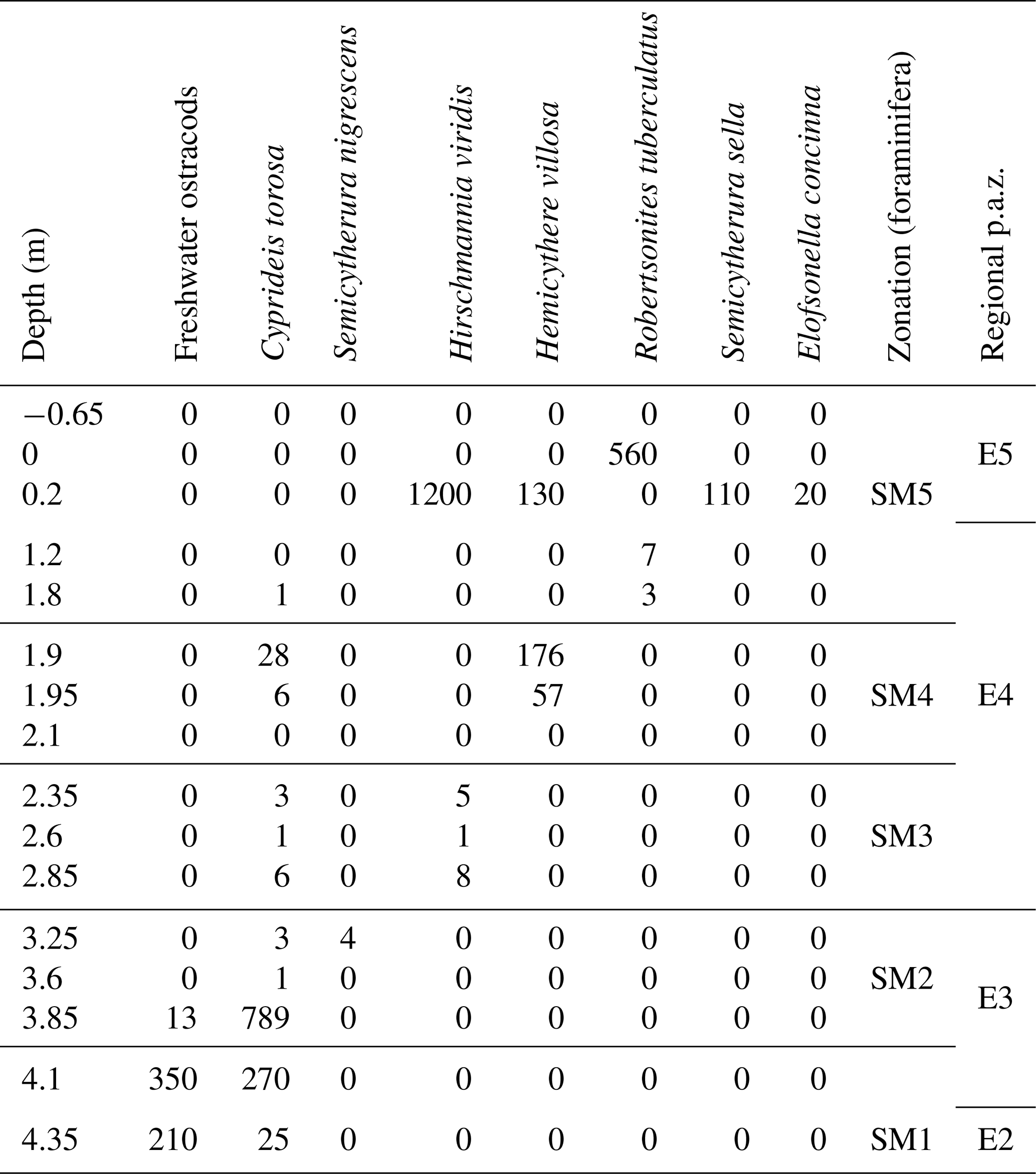
Foraminifera and ostracod assemblages have been studied in 22 samples from the Stensig section (Fig. 6; Table 2). The lowermost part of the studied succession contains an early interglacial freshwater ostracod assemblage, i.e. Metacypris cordata. The immigration of Cyprideis torosa may indicate an initial marine influx into the freshwater basin (Athersuch et al., 1989), but there is no foraminifera (SM1; Table 2). The first brackish-water foraminiferal species Elphidium albiumbilicatum (see Alve and Murray, 1999) is found in zone SM2, where it becomes totally dominant. This is at the transition between the Betula–Pinus zone (E2) and the Quercus–Fraxinus zone (E3). The immigration of Haynesina orbiculare, Elphidium selseyensis and Buccella frigida indicates gradually higher salinity and deeper waters during the deposition of zone SM3 (Murray, 1991, 2006), and a succeeding increase in Haynesina nivea and Elphidium balticum (previously referred to Elphidium incertum Williamson; see Knudsen et al., 2024) points to almost fully marine conditions with salinities higher than 25 in SM5 (see Lutze, 1965, 1974). This interpretation is supported by the ostracod assemblage. For instance, the species Robertsonites tuberculatus is an indicator of sublittoral conditions, living today in the deeper parts (18–32 m) of the south-western Baltic region at a relatively high salinity (Rosenfeld, 1977). A short-term reversal to subtidal brackish-water conditions is, however, indicated by high frequencies of the foraminiferal species Elphidium gunteri in zone SM4 (see Haake, 1962). It is still unclear whether this is due to some local environmental change or if it is rather a regional phenomenon. Marine conditions prevail at the Stensig section until the early part of the Picea zone (E5).
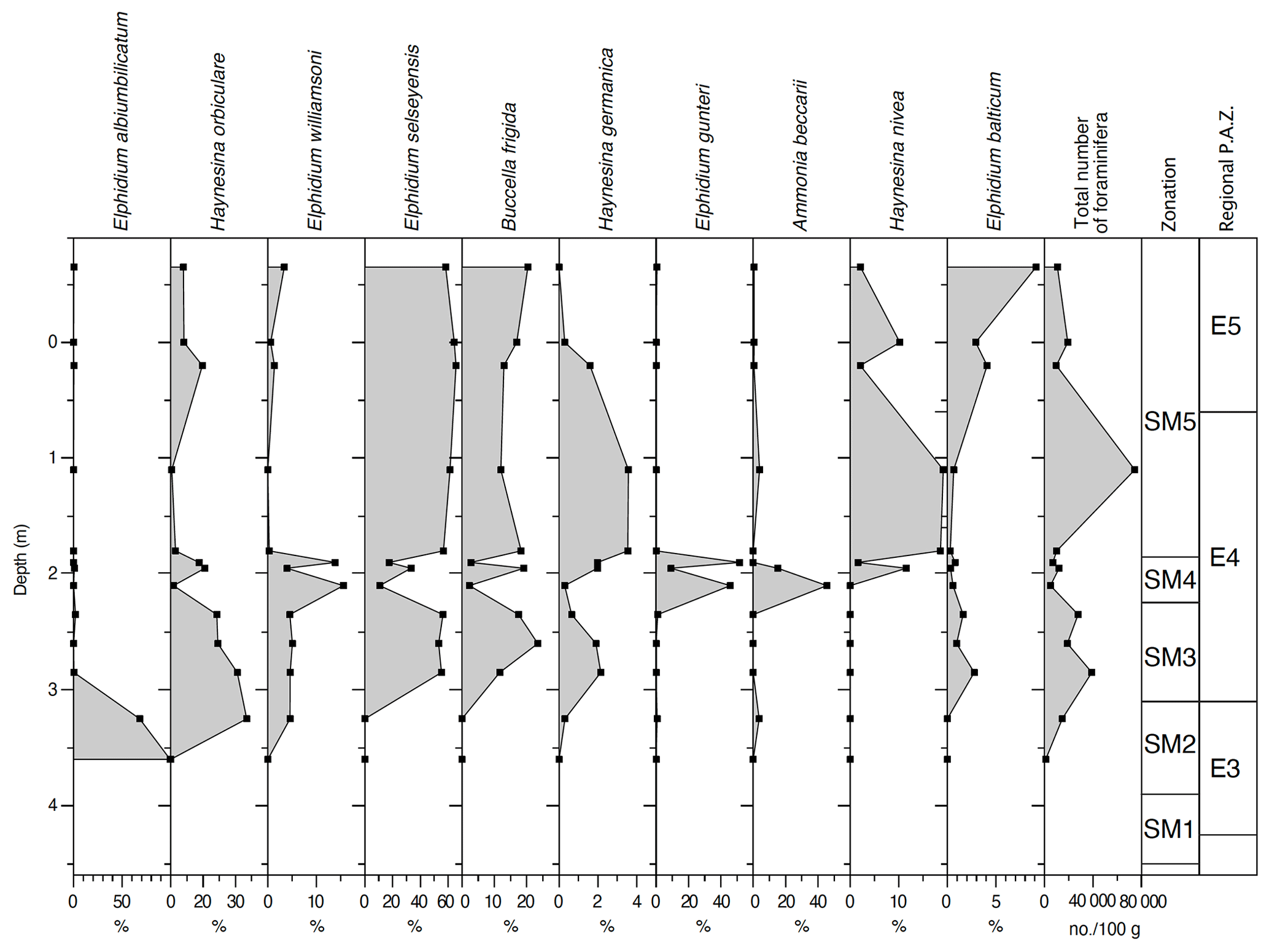
Figure 6Percentage distribution of the most important foraminiferal species, as well as the total number of specimens per 100 g of sediment in the Stensig section at Gammelmarke, southern Denmark. The combined foraminiferal and ostracod zonation (see also Table 1) is correlated with the regional pollen assemblage zones (Andersen, 1961, 1975).
Table 2Total number of freshwater ostracods and seven species of marine ostracods per 100 g of sediment in samples from the Stensig section at the Gammelmarke coast cliff, southern Denmark (combined foraminiferal and ostracod zonation; Figs. 6, 7), with correlation to the regional pollen assemblage zones (Andersen, 1961, 1975).
3.2.3 Summary
The correlation of the local pollen assemblage zones with the local foraminiferal zones and the regional pollen zones (Fig. 7) indicates that the sequence at the Stensig section represents Eemian regional pollen zones E1 to E5, based upon Andersen's later scheme (1961, 1975) for the Danish parastratotype sequence at Hollerup (Björck et al., 2000).
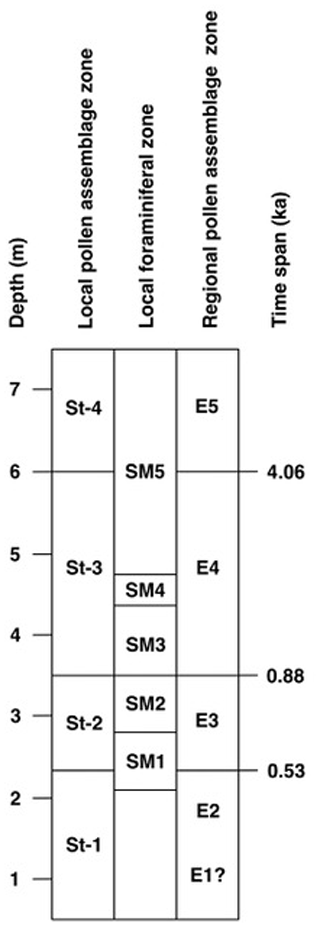
Figure 7Correlation of local pollen assemblage zones and the local foraminiferal zones of the Stensig section with the regional pollen zones of Andersen (1961, 1975) and the indication of ages for the pollen zone boundaries (Lauterbach et al., 2024).
3.3 Stohl
3.3.1 Lithology
The Stohl site is situated at the Baltic coast in northern Germany (Figs. 1, 2; 54°28′34.28′′ N, 10°9′14.87′′ E). The section at Stohl, which is exposed in a coastal outcrop, consists of a 6.50 m thick series of dislocated glacial and interglacial deposits. The lithostratigraphy of the Stohl site is shown in Fig. 8 (from Kubisch and Schönfeld, 1985).
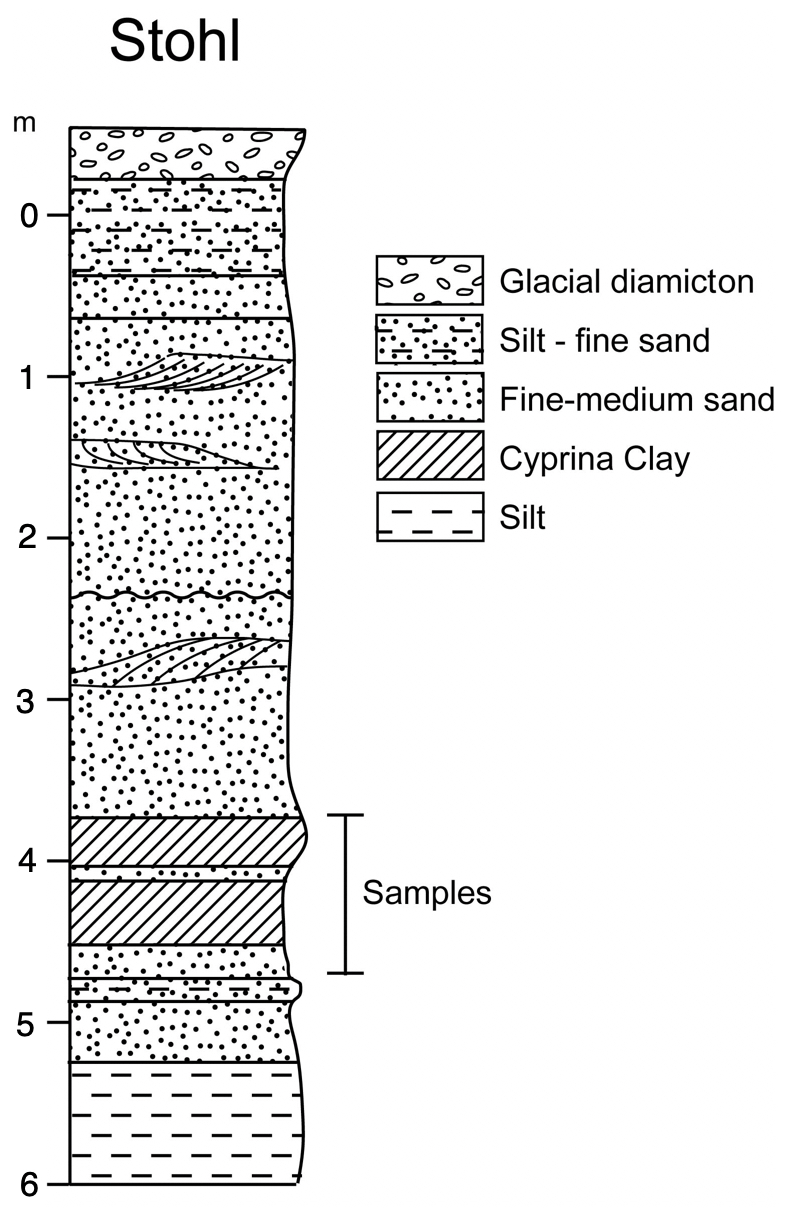
Figure 8Lithology of the Stohl section with an indication of the sampling interval of the Cyprina clay (from Kubisch and Schönfeld, 1985).
At the time of the sampling, it was not possible to access the interglacial deposits in the cliff due to a high sea level. Therefore, a coring was carried out on the beach using handheld drilling equipment. A series of samples were obtained from the 45 cm thick Cyprina clay and the underlying 30 cm of sandy shell-bearing sediment.
3.3.2 Pollen and spores
The samples collected for foraminiferal and ostracod analysis were subsampled for pollen analysis. The pollen and spores were generally well preserved in all samples. The sequence (Fig. 9) can be subdivided into three local pollen assemblage zones (l.p.a.z.'s), termed ST-1 to ST-3 (from base to top).

Figure 9Summary pollen diagram from the Stohl section, north Germany, with correlation to the regional pollen zones (Andersen, 1961, 1975).
The pollen sequence begins with an assemblage dominated by Pinus and Betula, with very low proportions of non-arboreal pollen. Filicales (Pteropsida) spores are very common, their presence being interpreted as representing local damp understorey environments, as noted above. This pre-temperate assemblage is replaced in zone ST-2 by an assemblage representing mixed-oak woodland, marked by an increase in Quercus and the appearance in the sequence of Alnus. Importantly, the expansion in oak is accompanied by Ulmus and other taxa typical of temperate mixed-oak forest. The uppermost part of the sequence records an increase in Corylus and corresponding decline in Quercus proportions. It is important to note, however, that the distribution of Quercus in the vegetation would have remained at similar levels as the previous zone, with Corylus occurring principally as an understorey shrub. The observed decline in Quercus proportions is mostly a result of the closed dataset, as evidenced by the concentration data. The sequence contains very low proportions of non-arboreal pollen throughout, indicating that the environment at the time of deposition was heavily forested.
The pollen sequence at Stohl can be closely correlated with the Eemian Stage, e.g. Herning (Jessen and Milthers, 1928), Hollerup (Andersen, 1961, 1975; Björck et al., 2000), Bispingen (Müller, 1974; Lauterbach et al., 2024), Ristinge Klint (Kristensen et al., 2000) and locations described by Zagwijn (1983, 1996) (Fig. 3). The Stohl diagrams can be zoned directly according to the scheme of Jessen and Milthers (1928) that was developed in Denmark and north-west Germany. Equally the succession can be zoned using Andersen's (1961, 1975) later scheme for the Danish Eemian parastratotype locality at Hollerup (Fig. 1; see also Björck et al., 2000). This correlation is based upon the l.p.a.z.'s defined here that are equated to r.p.a.z.'s as follows: ST-1 = E1/2, ST-2 = E3, St-3 = E4 (see Andersen, 1975; Zagwijn, 1983, 1996).
3.3.3 Foraminifera and ostracods
A total of 12 samples were processed and analysed for foraminifera and ostracods (Fig. 10). The Stohl record is characterized by an initial brackish-water phase, probably deposited in a near-coastal environment with Elphidium albiumbilicatum and Ammonia beccarii as the dominant species (zone S1) at salinity lower than 20 (see Alve and Murray, 1999; Murray, 1991, 2006). This shallow-water phase is followed by a gradual change to deeper waters and higher-salinity environments (20–25), indicated by a decrease in the two former shallow-water species and an increase specifically in Haynesina orbiculare and Elphidium selseyensis (zone S2) (see Lutze, 1965, 1974). Only one ostracod species was found at Stohl, i.e. the brackish-marine Cyprideis torosa. The correlation with the pollen stratigraphy shows that the marine ingression at Stohl took place at the very beginning of the Eemian, which is at the transition to pollen zone E3, and it presumably only prevailed a few hundred years into pollen zone E4.
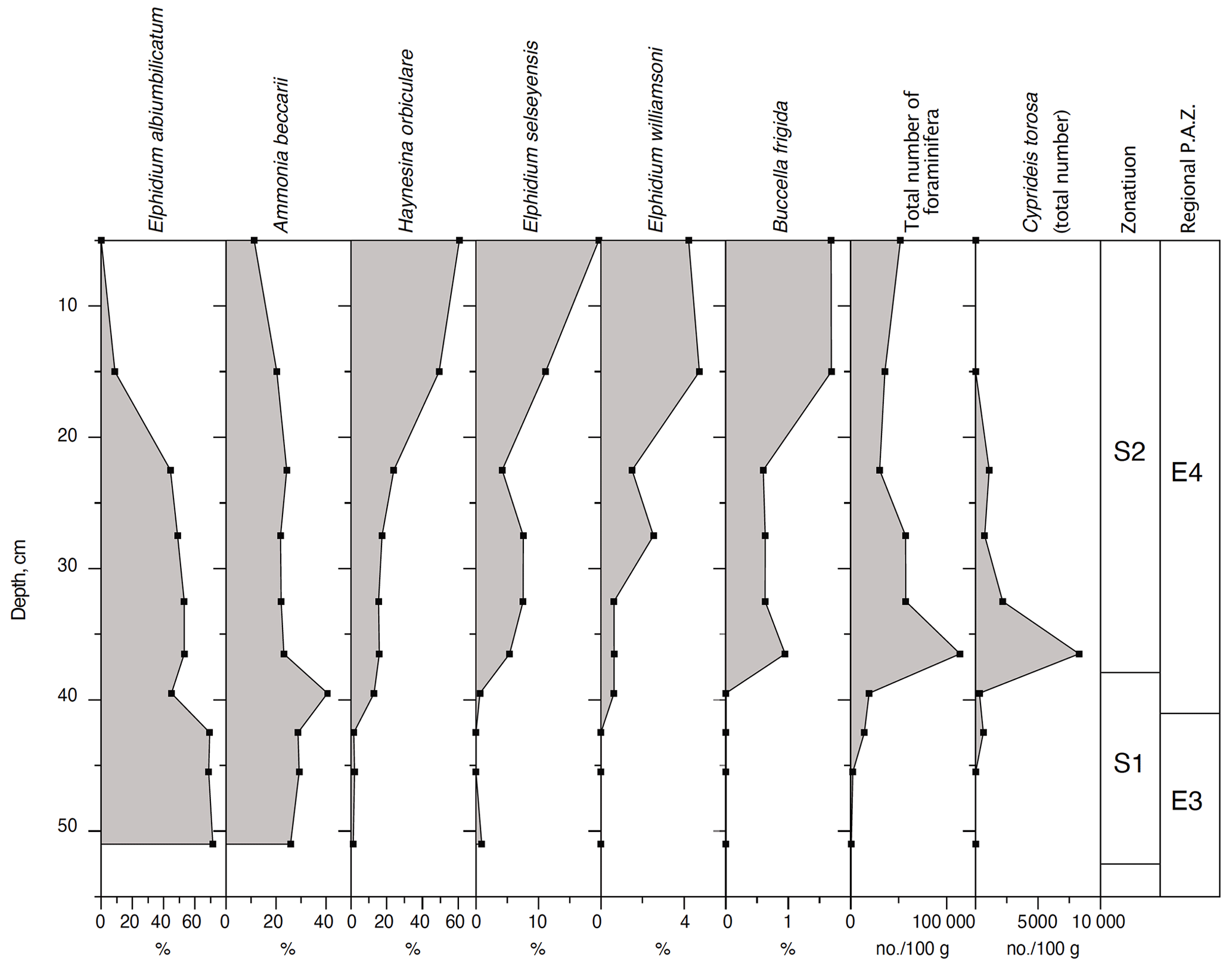
Figure 10Percentage distribution of the most important foraminiferal species and the total number of foraminiferal specimens per 100 g of sediment, as well as the number of the ostracod Cyprideis torosa per 100 g of sediment in the Stohl section, north Germany. The combined foraminiferal and ostracod zonation is correlated with the regional pollen assemblage zones (Andersen, 1961, 1975).
3.3.4 Amino-acid measurements
A total of 13 amino-acid measurements from Stohl have been carried out on the foraminiferal species Elphidium selseyensis. The mean value at Stohl was 0.08, i.e. close to the mean values obtained from Eemian deposits at Ristinge Klint (mean = 0.067 on E. selseyensis) and Mommark (mean = 0.07 on E. selseyensis) (BALTEEM participants, 2002; see also method and references in Knudsen and Sejrup, 1988).
3.3.5 Summary
As demonstrated above, the Stohl sequence spans Eemian regional pollen zones E1 to E4. Equally the succession can be zoned using Andersen's (1961, 1975) later scheme for the Danish lacustrine Eemian parastratotype locality at Hollerup (Fig. 1; see also Björck et al., 2000). The correlation of the local pollen assemblage zones with the local foraminiferal zones and the regional pollen zones of Andersen (1961, 1975), as well as indication of ages for the pollen zone boundaries, is shown in Fig. 11.
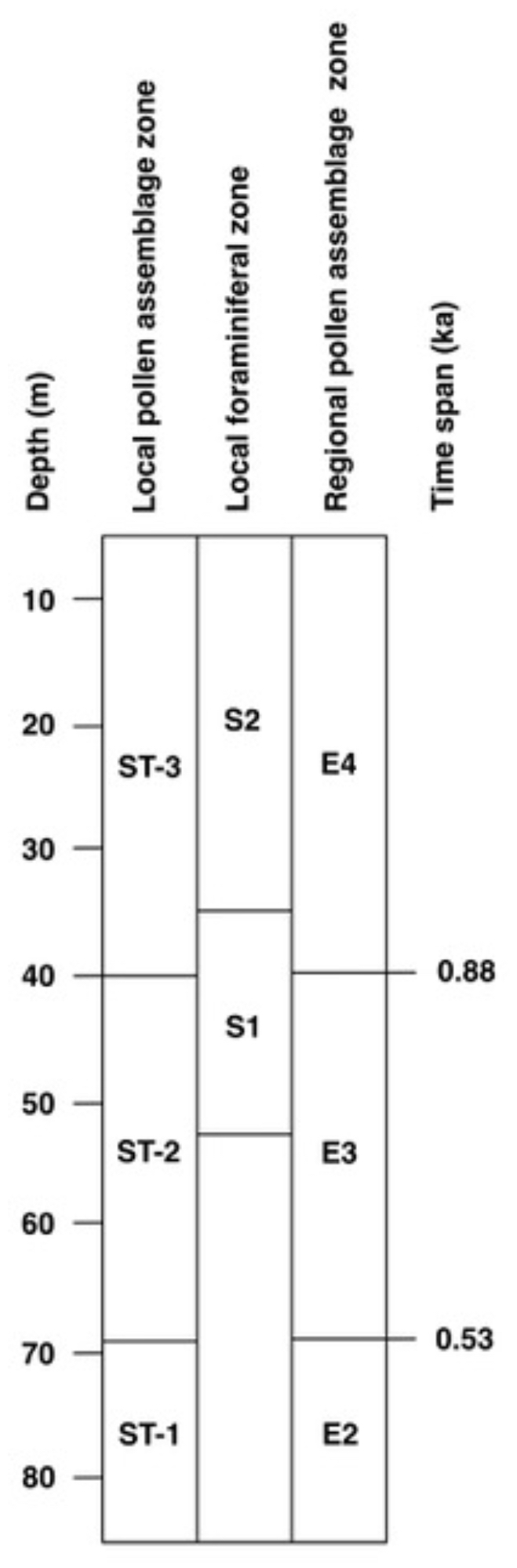
Figure 11Correlation of local pollen assemblage zones and local foraminiferal zones of the Stohl section with the regional pollen zones of Andersen (1961, 1975), as well as an indication of ages for the pollen zone boundaries (Lauterbach et al., 2024).
3.4 Vejsnæs Nakke
3.4.1 Lithology
The Vejsnæs Nakke site was originally identified by Jessen and Milthers (1928). Here the Cyprina clay is exposed in the cliff headland on the southern point of the small island of Ærø, south of Fyn (Figs. 1, 2; 54°48′59.50′′ N, 10°25′26.84′′ E). The unit occurs as a glacially thrust inclusion block within till. It is 30 cm thick and is cut through by shear and fault planes. The deposit is light grey clay resting on a unit of “freshwater” sand. Samples were taken for pollen analysis, and block samples were collected for molluscs by Svend Funder. No samples were analysed for foraminifera and ostracods from this locality.
3.4.2 Pollen and spores
A total of eight samples were collected from the Cyprina clay at Vejsnæs Nakke for fossil pollen and spore analysis (Fig. 12). The sequence can be subdivided into two local pollen assemblage zones (l.p.a.z.'s) on the basis of the visual recognition of significant changes in the sequence. The lowermost zone (V1) is dominated by high proportions of Pinus and Betula pollen, with the only significant herbaceous pollen being Poaceae and Cyperaceae. Relatively high proportions of pre-Pleistocene palynomorphs suggest the inwash of reworked material via streams entering the basin and possibly also local coastal erosion. The lowermost part of the assemblage is typical of that found surrounding freshwater bodies, with a local fringing vegetation of wetland, marsh and aquatic plants, with mixed pine and birch woodland on drier soils.
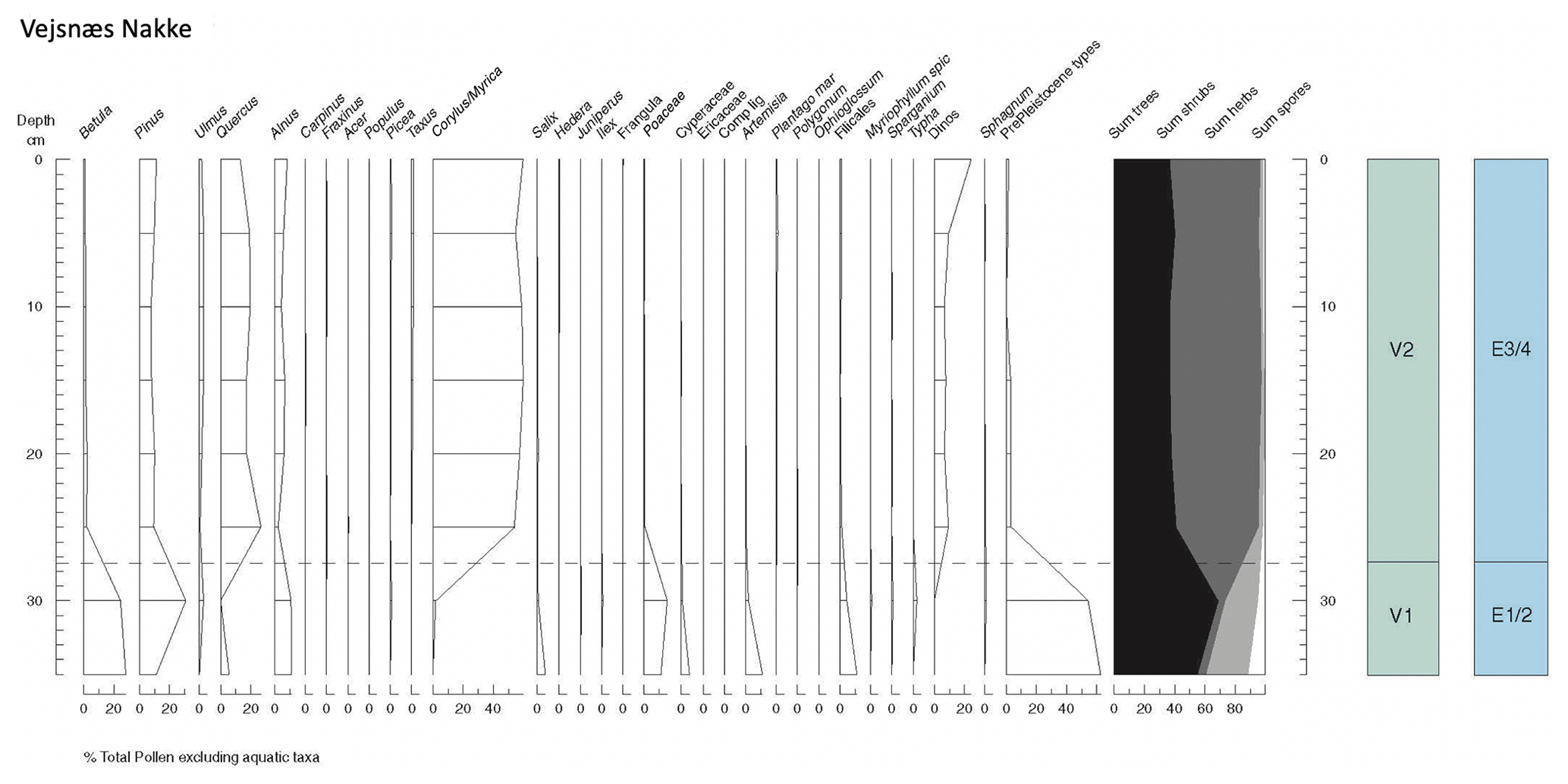
Figure 12Summary pollen diagram from the Vejsnæs Nakke section on the island of Ærø, southern Denmark, with correlation to the regional pollen zones (Andersen, 1961, 1975).
This assemblage is succeeded in zone V2 by an expansion of Corylus and Quercus pollen, with a continuous curve of Taxus in the upper part. The vegetation at this point is fully temperate mixed-oak woodland with warmth-demanding taxa such as Hedera.
The pollen sequence at Vejsnæs Nakke can also be closely correlated with those of the Eemian Stage, e.g. Herning (Jessen and Milthers, 1928), Bispingen (Müller, 1974; Lauterbach et al., 2024), Ristinge Klint (Kristensen et al., 2000) and locations described by Zagwijn (1983, 1996). On the basis of the l.p.a.z.'s defined here, they are equated to r.p.a.z.'s as follows: V1 = E1/E2, V2 = E3/E4 (see Andersen, 1975; Zagwijn, 1983).
The comparison with sequences such as Mommark (Gibbard and Glaister, 2006) and Hollerup (Andersen, 1965; Björck et al., 2000), as well as nearby terrestrial lacustrine localities, unequivocally demonstrates that there is a hiatus in the sequence at 27–28 cm. This hiatus causes the Quercus zone (i.e. r.p.a.z. E2) to be incompletely unrepresented at this site. This discontinuous sequence arises from the fact that the sediments are highly disturbed by subsequent glaciotectonics (as noted above). It is therefore likely that the absent sediments were removed by faulting, these structures being avoided during sampling.
3.4.3 Summary
Taken together the evidence from Vejsnæs Nakke indicates that the sediments are of Eemian age, and the diagrams can be zoned directly according to the scheme of Jessen and Milthers (1928) that was developed in Denmark and north-west Germany. Equally the succession can be zoned using Andersen's (1961, 1975) later scheme for the Danish lacustrine Eemian parastratotype locality at Hollerup (see also Björck et al., 2000). Indeed, the similarity between the tree pollen curves at Hollerup, 180 km to the north-north-west, and those at Stohl is striking and further reinforces the correlation. On this basis the Vejsnæs Nakke sediments represent regional pollen zones E1/E2 to E3/E4 (see Andersen, 1975; Zagwijn, 1983).
Having established the time represented by the Eemian sediments and the marine intervals at each of the investigated localities, it is now possible to compare and correlate the individual sequences with each other and previously studied sites within the region. The study of the benthic foraminifera and ostracods indicates bottom-water conditions during the Eemian Stage interglacial. In addition, the establishment of a chronology for each site provides a foundation for the study of specific time slices or short time intervals over a larger area.
As noted above, the palynological correlation of the marine intervals in each site of the south Baltic region with the Bispingen record in northern Germany, the latter originally based on that proposed by Müller (1974), is here related to the revised chronology of Lauterbach et al. (2024). This provides a relative chronology for the sites shown in Fig. 13. Salinity interpretations for the records are indicated by the colour scale on the plot, which extends from the North Sea coast in the west through a marine Eemian “fjord system” across northern Germany to the southern Danish sites.
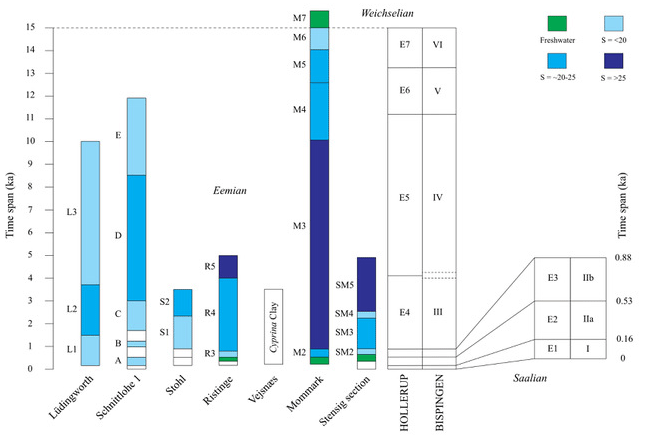
Figure 13Correlation of Eemian records from the North Sea coast in the west through northern Germany to the study area in the western Baltic. The diagram shows the duration of the Eemian record at each site according to the pollen zonation, as well as the timing of the marine environment at each site (salinity (S) indications are shown in graded blue colours). The foraminiferal zonation for each record is given on the left-hand side of each column. The time span refers to the Lauterbach et al. (2024) stratigraphy.
The Eemian foraminiferal assemblage zonal boundaries, obtained from previous studies of cores from Lüdingworth (Knudsen, 1988; see below) and Schnittlohe 1 (Knudsen, 1985; see below) in Schleswig-Holstein, northern Germany, have been re-interpreted in addition based upon the new age model. These two sites are representative of sediment deposition during a possible period of marine connection across northern Germany, in the Kiel Canal area (Fig. 1), in part of the interglacial (Kosack and Lange, 1985). On the basis of a correlation of the pollen stratigraphy from the two sites with that from Bispingen, the foraminiferal results are now entered into the floating timescale (Fig. 13).
4.1 Lüdingworth
Lüdingworth is located close to the mouth of the Elbe River on the North Sea coast of Germany (Figs. 1, 2; 53°47′54.68′′ N, 8°45′36.85′′ E). Here, Eemian deposits are found between 19 to 14 m depth in the borehole (i.e. 18–13 m below sea level), being both under- and overlain by glaciogenic sediments (Knudsen, 1988). The marine deposits occur immediately overlying a thin peat stratum. The environment was initially shallow and brackish, indicated by extremely high frequencies of Elphidium albiumbilicatum (see Knudsen, 1988: their Fig. 4, zone L1, at the base of r.p.a.z. E2). This was rapidly superseded by a period with more saline, sublittoral conditions, indicated by additional species such as Elphidium selseyensis, E. williamsoni and Buccella frigida (zone L2). The final marine stage (zone L3) records a return to shallow and brackish waters caused by a fall in relative sea level. This is indicated by a high dominance of the species Ammonia beccarii. Salinity interpretations, foraminiferal zonation and the pollen stratigraphy are shown in Fig. 13.
4.2 Schnittlohe 1
Schnittlohe 1 is located further towards the east in the central part of Schleswig-Holstein, northern Germany (Figs. 1, 2; 54°8′27.85′′ N, 9°23′19.22′′ E). At this locality Eemian Stage deposits occur between 17.3 and 13.0 m depth in the borehole (ground level is 0.30 m above sea level). Here the foraminifera indicate shallow-water conditions, particularly early in the sequence when minor marine inundations occurred (zones A, B and C, i.e. in r.p.a.z.'s E2 and E4; Fig. 13). This is indicated by high percentages of Ammonia beccarii and E. albiumbilicatum together with E. gunteri. The first ingression in the area might have occurred as early as the base of regional pollen zone E2. The succeeding immigration of species such as Elphidium balticum, Buccella frigida, Haynesina nivea and H. orbiculare in foraminiferal zone D (i.e. pollen zones E4–E5) indicates the establishment of more persistent open-marine conditions and water depths of up to 10–20 m. Evidence of withdrawal of the Eemian Baltic Sea from the area, indicated by the dominance of Haynesina germanica and A. beccarii, is recorded in the upper part of the marine succession (zone E, i.e. late in r.p.a.z. E5 and early in E6).
4.3 Ristinge Klint
According to Kristensen et al. (2000) and Knudsen et al. (2011), the pollen sequence at Ristinge Klint (Figs. 1, 2), on the island of Langeland south-east of Fyn (54°49′41.44′′ N, 10°36′35.08′′ E), can be closely correlated with that of the Last Interglacial (Eemian Stage) of north-west Europe. The sediments represent Jessen and Milthers' (1928) pollen zones d–g or Andersen's (1961, 1975) zones E2–E5. The correlation with the revised Bispingen chronology demonstrates that the interglacial Ristinge Klint sequence spans ca. 5000 years. Here, the glacially thrust block sequence records a marine inundation entering the initial freshwater basin at the base of regional pollen zone E3. A total dominance of the foraminiferal species Elphidium albiumbilicatum indicates shallow, low-salinity conditions in the lower part (zone R3; Fig. 12; called SMR3 by Kristensen et al., 2000, and Knudsen et al., 2011). This is followed by an increase in e.g. Haynesina orbiculare and Buccella frigida, indicating increased salinity (20–25) and water depth in zone R4 and subsequently close to normal marine salinity in zone R5 at the top of the Cyprina clay. A similar environmental change is indicated by the ostracod assemblages.
If the initial marine ingression is taken to coincide with the base of the Cyprina clay at Ristinge Klint, marine conditions were established at ca. 530 years after the beginning of the interglacial, and a major change in salinity is observed at ca. 850 years after the beginning of the interglacial. This event may result from a western marine opening to the area through the Danish belts. Another major change in current activity is registered at the regional pollen zone boundary E4/E5, i.e. ca. 4060 years after the beginning of the stage, at the local pollen zone boundary between RK III and RK IV.
4.4 Mommark
The site at Mommark is located in the cliff on the east of the island of Als (Figs. 1, 2; 54°55′50.54′′ N, 10°2′39.83′′ E). Apart from the fact that the greater proportion of the earliest Eemian Betula r.p.a.z. (E1) is missing at Mommark (i.e. ca. 100 years), as well as by implication the Saalian–Eemian stage boundary, in total it appears that the sequence spans ca. 15 000 years or over 98 % of the interglacial time (Fig. 13). Mommark is the only site which preserves almost the entire Eemian event, representing the interglacial sea-level cycle, from freshwater initially to marine in the upper part of r.p.a.z. E2 at ∼ 400–500 years (in foraminiferal zone M2; called Mom 2 by Kristensen and Knudsen, 2006). A pronounced salinity increase occurs in the uppermost part of E3, continuing into E4 and E5 (foraminiferal zone M3), as indicated by both foraminifera (e.g. Buccella frigida and Haynesina orbiculare; Kristensen and Knudsen, 2006) and ostracods. Decreasing salinity occurs in foraminiferal zones M4–M5, as indicated by a marked increase in Elphidium selseyensis (i.e. the uppermost part of E5 to the lowermost part of E7). A return to shallow-water assemblages in foraminiferal zone M6, indicated by the appearance of Elphidium albiumbilicatum, is followed by freshwater conditions as shown by freshwater ostracods in the uppermost pollen assemblage zone E7. This potentially reflects local marine regression.
Assuming that the sequence is continuous through the interglacial at Mommark, the sedimentation rates vary considerably (calculated on the basis of current, i.e. post-compaction thicknesses). On this basis initial sedimentation was relatively rapid during r.p.a.z.'s E1 to E2, slowing in E3 to E4 and remaining low throughout the rest of the sequence. While it is unclear why the sedimentation rates were so slow, it is possible that intermittent deposition in shallow water, possibly disturbed by local currents and bioturbation, may have had a significant impact. The sedimentation rates indicate that sediment supply throughout the interglacial, particularly through the regional pollen assemblage zones E3 to E7, was restricted. This presumably reflects the fact that the streams feeding the basin were transporting relatively low concentrations of suspended-load clay and silt – a situation expected under conditions of undisturbed, dense forest vegetation that is generally accepted as having colonized the hinterland throughout the interglacial period.
These interglacial sediments at Mommark represent a glacially thrust slice which has been uplifted by glaciers from the sea floor just east of the island of Als at the east coast of Jylland (Figs. 1, 2). Seismic measurements show that a Quaternary valley (up to 200 m deep) is present in this area, a valley which is presumably filled partially by marine Eemian deposits (Eiríksson et al., 2006). This deep trench continues towards the north into the Lillebælt and may have represented a deep marine connection through to the Kattegat during the Eemian Stage (Kristensen et al., 2000).
The apparent uniformity of the vegetational sequence and its zones means that the chronology established can be applied with caution to the whole south-western Baltic area (e.g. Miller and Mangerud, 1985; Björck et al., 2000; Kristensen et al., 2000). The deposits in the south-western Baltic area, generally represented by the marine Cyprina clay, formed in inner-shelf, tidal-flat or fjord-like environments, frequently under conditions of hypoxia (see Funder and Balic-Zunic, 2006; Nielsen et al., 2007). The overlying Tapes sand at some of the sites represents a relatively shallower-water facies than the Cyprina clay.
As noted above, the first possible marine ingression at Schnittlohe 1 and Lüdingworth appears to have occurred as early as the base of regional pollen zone E2 (Menke, 1985; Funder et al., 2002), according to the zonation of Andersen (1965). This is earlier than that known from Stohl, Ristinge Klint, Mommark and the Stensig section, where the marine ingression occurred close to the base of r.p.a.z. E3. A comparison with marine Eemian sites on the west coast of Denmark shows that the first marine inundation was broadly simultaneous throughout the entire area, i.e. at the base of the Quercus r.p.a.z. (E3) (Winn et al., 2000; Konradi et al., 2005).
Notwithstanding the caveat noted earlier concerning the accuracy of the chronology, the marine regression, seen as a shallowing of the water depth, was initiated at ca. 12 500 years after the beginning of the Eemian Stage at Mommark (e.g. Funder and Balic-Zunic, 2006; Kristensen and Knudsen, 2006). Given that the interglacial ended ∼ 15 000 years ago, according to Lauterbach et al. (2024), this equates to the transition from marine to freshwater conditions at the site. Following the independent astronomical chronology proposed by Shackleton et al. (2003), which gives an age of 130 ka for the beginning of the Eemian, the full marine regression in the south-western part of the Baltic Sea began at 115.40 ka (see Sánchez Goñi et al., 2005). This date is close to that assigned to the deep-sea Marine Isotope Substage (MIS) 5e–5d boundary indicated by the δ18O benthic signal, suggesting that global sea-level fall coincided with that in the south-western Baltic region. It also implies that the main component of sea-level fall in the region is the global eustatic signal rather than the result of local isostatic adjustment of the Baltic Shield region. Evidence for the shallowing of water, possibly reflecting initial marine regression from the area, is found at Mommark late in regional pollen zone E5 and possibly at both Lüdingworth and Schnittlohe 1 in late E5 to early E6. This corresponds to the sea-level curve (Zagwijn, 1983, 1996) constructed from the Netherlands.
The ostracod assemblages show that a series of freshwater lakes existed in the area prior to the marine inundation. The variation in the timing of the initiation of sedimentation at the various sites, including the absence of the Betula zone r.p.a.z. E1, at some sites, potentially reflects the irregular morphology of the pre-interglacial topography, inherited from the immediately prior Late Saalian glacial and periglacial environment. Despite this variation, the various fossil assemblages studied in the region all support an interpretation of rapid relative sea-level rise at an early stage of the interglacial. Within a few hundred years, the conditions approached fully marine, with the highest salinities apparently established during regional pollen zone E4 and the lower part of E5 (Fig. 13).
A comparison of the foraminiferal assemblages and their stable isotopes with the surface water dwellers, dinoflagellates, indicates that there was stratification of the water masses, especially during the maximum transgression (Kristensen and Knudsen, 2006; Head, 2007). In general, all the fossil groups indicate Eemian temperatures some degrees higher than at present in the same area. The foraminifera also clearly show that temperatures were higher than during the Holocene. Likewise, the salinity was considerably higher during the Eemian high-sea-level stand than during that in the Holocene.
Recent studies of the geochemistry of foraminiferal tests from a series of Eemian sites in the western Baltic region (Ni et al., 2021) support a previous interpretation of bottom-water temperature and salinity in the area being higher than in recent (i.e. modern) times. At Ristinge, these authors found a clear increase in both temperature and salinity from pollen zone E3 through E4 and into E5. In addition, Ni et al. (2021) demonstrated that greater seasonal temperature differences during the first half of the interglacial were mainly caused by higher summer temperatures rather than lower spring temperatures. This was ascribed to the increase in summer insolation and potentially a reduced spring insolation. These results support previously published evidence for increased seasonality during the Last Interglacial compared to today in mid- to high latitudes (see Berger and Loutre, 1991; Laskar et al., 2004; Bova et al., 2021). On the basis of modelling studies, Ni et al. (2023) further discussed how the substantial changes in seasonal insolation may have affected the hydrography and water-mass exchange in the North Sea and Baltic Sea regions.
The important Eemian-aged marine gateway from the North Sea via the Kattegat into the Baltic was the deep trench through northern Denmark, which may have had an open connection through the Alnarp Valley (Fig. 1) towards the south-east (Funder et al., 2002). Eemian deposits from this trench have been described from several cores in northern Jylland and Kattegat, including Skærumhede I (Knudsen and Lykke-Andersen, 1982), Apholm (Knudsen, 1984) and Anholt (Seidenkrantz, 1993a, b).
The markedly limited thicknesses of the marine sediments in the south-west Baltic area indicate that sediment supply throughout the interglacial, particularly through the densely forested regional pollen zones E3–E6, was restricted. Although it is generally considered that undisturbed, dense forest vegetation colonized the hinterland throughout the interglacial period, the implication is that the assumed limited forest cover on the adjacent land areas, proposed following vegetational modelling by Pearce et al. (2023), is not supported by the depositional evidence (see Roebroeks et al., 2024).
Marine Eemian deposits occur widely in the western Baltic area and are often exposed as glacially dislocated sediments in coastal cliffs in the area. The geographical location of the area close to the Danish belts with marine connections to the Kattegat, as well as to the Eemian marine connection across northern Germany, makes it a key area for the understanding of the regional palaeoenvironmental changes through the interglacial. The sites discussed here represent the Last Interglacial Eemian Stage, an interpretation which is based on their pollen content and comparison with regional pollen zonation schemes.
The establishment of a chronology for each site has provided a firm framework. Palynological correlation of the marine and non-marine intervals with the Bispingen record in northern Germany gives an independent chronology for correlation and comparison of the local depositional sequences, including the timing and effects of the marine transgression and regression in the south-western Baltic during the interglacial.
The study of a series of critical sequences from the area, including benthic foraminifera and ostracods, has demonstrated the bottom-water conditions during the Eemian Stage. The ostracod assemblages show that freshwater lakes existed in the south-western Baltic basin prior to the marine inundation. The earliest marine transgression in the south-west at Lüdingworth and Schnittlohe in northern Germany appears to have occurred at the base of pollen zone E2, whilst at Ristinge Klint, the Stensig section, Mommark and Stohl the marine ingression occurred at the base of regional pollen zone E3. Fully marine conditions occurred at the base of r.p.a.z. E4, continuing into the early part of E5.
The linking of the northern German sequences at Lüdingworth and Schnittlohe 1 to those in the south-west Baltic demonstrates that the transgression occurred both through the Danish belts and through the Kiel Canal area in Schleswig-Holstein (Fig. 1), the results indicating that a connection with the North Sea was present in the area during the interglacial sea-level maximum highstand interval.
Evidence for the shallowing of water, potentially reflecting initial marine regression from the area, is found at Mommark late in regional pollen zone E5 at ca. 12 500 years after the beginning of the stage. This compares favourably to that at both Lüdingworth and Schnittlohe 1 late in zone E5 to early zone E6 (i.e. ca. 12 000 years). The regression of the sea during zone E6 was followed by freshwater deposition in zone E7 at Mommark in the south-west Baltic area. This timing compares favourably with the sea-level record from the Netherlands and that from the deep-sea sequences. Further it contributes to the detailed impact of sea-level and climatic change, which provides a vital analogue towards understanding how these processes might be expected to operate during the present interglacial.
Palaeontological data are available from the authors.
The project, the analyses, the interpretation and the writing of the article were undertaken equally by both authors. Both authors have read and agreed on the published version of the paper.
The contact author has declared that neither of the authors has any competing interests.
Publisher’s note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors.
The authors thank Peter Kristensen, Jón Eiríksson, Chris Glaister and Svend Funder for field assistance and Steve Boreham and Peter Kristensen for laboratory preparation. We also thank Peter Kristensen for the analyses of foraminifera and ostracods; Chris Glaister for assistance with the pollen analyses; and Jón Eirikson, Philip Stickler, Sebastian Gibson and Liliia Hebryn-Baidy for help with drafting the figures. We warmly thank our fellow BALTEEM participants for discussions and additional analyses. Finally, we express our gratitude to reviewer Brigitte Urban, an anonymous reviewer and the editor Elisabeth Dietze for their suggestions and observations that improved the paper.
This research has been supported by the European Commission, Fourth Framework Programme (grant no. ENV4-CT98-0809).
This paper was edited by Elisabeth Dietze and reviewed by Brigitte Urban and one anonymous referee.
Aalbersberg, G. and Litt, T.: Multiproxy climate reconstructions for the Eemian and Early Weichselian, J. Quaternary Sci., 13, 367–390, 1998.
Alve, E. and Murray, J. W.: Marginal marine environments of the Skagerrak and Kattegat: a baseline study of living (stained) benthic foraminiferal ecology, Palaeogeogr. Palaeocl., 146, 171–193, 1999.
Andersen, S. T.: Vegetation and its environment in Denmark in the Early Weichselian Glacial (Last Glacial), Danmarks Geologiske Undersøgelse II. Række, 75, 1–175, 1961.
Andersen, S. T.: Interglacial plant successions in the light of environmental changes, in: Report of the VIth International Congress on the Quaternary, Warsaw 1961, II, International Association for Quaternary Research, 13 pp., 1964.
Andersen, S. T.: Interglacialer og interstadialer i Danmarks Kvartær, Meddelelser fra Dansk Geologisk Forening, 15, 486–506, 1965.
Andersen, S. T.: Interglacial vegetational succession and lake development in Denmark, Palaeobotanist, 15, 117–127, 1966.
Andersen, S. T.: The Eemian freshwater deposit at Egernsund, South Jylland, and the Eemian landscape development in Denmark, Danm. Geol. Undersog. (DGU) Årbog 1974, 49–70, https://doi.org/10.22008/gpub/38240, 1975.
Andrew, R.: A Practical Pollen Guide to the British Flora, Quaternary Research Association, Cambridge, 139 pp., ISBN 0907780032, 9780907780038, 1984.
Athersuch, J., Horne, D. J., and Whitaker, J. E.: Marine and brackish water ostracods, in: Synopsis of the British Fauna (n.s.), Brill, Leiden, 43, 343 pp., 1989.
BALTEEM participants: Palaeoenvironmental and Palaeoclimatic Evolution of the Baltic Sea Basin During the Last Interglacial (Eemian, Mikulino), Final report: October 2002, Contract no: ENV4-CT98-0809 BALTEEM, 180 pp., 2002.
Bates, C. D., Coxon, P., and Gibbard, P. L.: A new method for preparation of clay-rich sediment samples for palynological investigations, New Phytol., 81, 459–463, 1978.
Berger, A. and Loutre, M.-F.: Insolation values for the climate of the last 10 million years, Quaternary Sci. Rev., 10, 297–317, 1991.
Birks, H. J. B.: Past and present vegetation of the Isle of Skye. A palaeoecological study, Cambridge University Press, Cambridge, 415 pp., ISBN-13.978-0521085335, 1973.
Björck, S., Noe-Nygaard, N., Wolin, J., Houmark-Nielsen, M., Hansen, H. J., and Snowball, I.: Eemian Lake development, hydrology and climate – a multi-stratigraphic study of the Hollerup site in Denmark, Quaternary Sci. Rev., 19, 509–536, 2000.
Bova, S., Rosenthal, Y., Liu, Z., Godad, S. P., and Yan, M.: Seasonal origin of the thermal maxima at the Holocene and the last interglacial, Nature, 589, 548–553, 2021.
Caspers, G.: Die eem- und weichselzeitliche Hohlform von Groß Todtshorn (Kr. Harburg; Niedersachsen) – Geologische und palynologische Untersuchungen zu Vegetation und Klimaverlauf der letzten Kaltzeit, Schriftenreihe, Zeitschrift der Deutschen Geologischen Gesellschaft, 4, 7–59, 1997.
Caspers, G., Merkt, J., Müller, H., and Freund, H.: The Eemian interglaciation in northwestern Germany, Quaternary Res., 58, 49–52, 2002.
CLIMAP project members: The Last Interglacial Ocean, Quaternary Res., 21, 123–224, 1984.
Cohen, K. M., Cartelle, V., Barnett, R., Busschers, F. S., and Barlow, N. L. M.: Last Interglacial sea-level data points from Northwest Europe, Earth Syst. Sci. Data, 14, 2895–2937, https://doi.org/10.5194/essd-14-2895-2022, 2022.
Ditlefsen, C.: Luminescence Dating of Danish Quaternary Sediments, unpublished PhD thesis, Department of Earth Science, Aarhus University, 154 pp., 1991.
Donner, J.: The Quaternary History of Scandinavia, in: World and Regional Geology Series, Vol. 7, Cambridge University Press, 200 pp., https://doi.org/10.1017/S0016756800007639, 1995.
Eiríksson, J., Kristensen, P. H., Lykke-Andersen, H., Brooks, K., Murray, A., Knudsen, K. L., and Glaister, C.: A sedimentary record from a deep Quaternary valley in the southern Lillebælt area, Denmark: Eemian and Early Weichselian lithology and chronology at Mommark, Boreas, 35, 320–331, 2006.
Ellis, B. F. and Messina, A.: Catalogue of Foraminifera (with Supplements), American Museum of Natural History and Micropaleontology Press, New York, https://foraminifera.eu/index.html (last access: 8 November 2024), 1949.
Feyling-Hanssen, R. W., Jørgensen, J. A., Knudsen, K. L., and Lykke-Andersen, A.-L.: Late Quaternary Foraminifera from Vendsyssel, Denmark and Sandnes, Norway, B. Geol. Soc. Denmark, 21, 67–317, 1971.
Forchhammer, G.: Oversigt over det Kongelige Danske Videnskabernes Selskabs Forhandlinger for 1841, 63–66, 1842.
Funder, S. and Balic-Zunic, T.: Hypoxia in the Eemian: mollusc faunas and sediment mineralogy from Cyprina Clay in the southern Baltic region, Boreas, 35, 367–377, 2006.
Funder, S., Demidov, I., and Yelovicheva, Y.: Hydrography and mollusc faunas of the Baltic and the White Sea-North Sea seaway in the Eemian, Palaeogeogr. Palaeocl., 184, 275–304, 2002.
Gibbard, P. L. and Glaister, C.: Pollen stratigraphy of the Late Pleistocene sediments at Mommark, Als, South Denmark, Boreas, 35, 332–348, 2006.
Glaister, C. G. and Gibbard, P. L.: Pollen stratigraphy of the Late Pleistocene marine sediments at Nørre Lyngby and Skagen, North Denmark, Quaternary Sci. Rev., 17, 839–854, 1998.
Gottsche, C.: Über den Tapes-Sand von Steensigmoos, Zeitschrift der Deutschen Geologischen Gesellschaft, 56, 181–184, 1904.
Gripp, K.: Erdgeschichte von Schleswig-Holstein, Karl Wachholtz, Neumünster, 311 pp., ISBN-13. 978-3529053306, 1964.
Haake, F. W.: Unterzuchungen an der Foraminiferen-Fauna im Wattgebiet zwischen Langeoog und dem Festland, Meyniana, 12, 25–64, 1962.
Hahne, H., Kemle, S., Merkt, J., and Meyer, K.-D.: Eem-, weichsel- und saalezeitliche Ablagerungen der Bohrung “Quakenbrück GE 2”, Geologisches Jahrbuch, Reihe A, 134, 9–69, 1994.
Head, M.: Last Interglacial (Eemian) hydrographic conditions in the southwestern Baltic Sea based on dinoflagellate cysts from Ristinge Klint, Denmark, Geol. Mag., 144, 987–1013, 2007.
Hinsch, W.: Die Molluskenfauna des Eem-Interglazials von Offenbüttel-Schnittlohe (Nord-Ostsee-Kanal, Westholstein), Geologisches Jahrbuch, Reihe A, 86, 49–62, 1985.
Houmark-Nielsen, M.: Pleistocene stratigraphy and glacial history of the central part of Denmark, B. Geol. Soc. Denmark, 36, 1–189, 1987.
Jessen, K. and Milthers, V.: Stratigraphical and palaeontological studies of interglacial freshwater deposits in Jutland and northwest Germany, Danmarks Geologiske Undersøgelse II. Række, 48, 1–379, 1928.
Knudsen, K. L.: Foraminiferal stratigraphy in a marine Eemian-Weichselian sequence at Apholm, North Jutland, Bull. Geol. Soc. Denmark, 32, 169–180, 1984.
Knudsen, K. L.: Foraminiferal faunas in Eemian depostis of the Oldenbüttel area near the Kiel Canal, Germany, Geologisches Jahrbuch, Reihe A, 86, 27–47, 1985.
Knudsen, K. L.: Marine Interglacial Deposits in the Cuxhaven Area, NW Germany: A Comparison of Holsteinian, Eemian and Holocene Foraminiferal Faunas, E&G Quaternary Sci. J., 38, 69–77, https://doi.org/10.3285/eg.38.1.06, 1988.
Knudsen, K. L.: A long marine Eemian-Weichselian shelf record in North Denmark, Scandinavia, in: Start of a Glacial, edited by: Kukla, G. and Went, E., Proceedings of the Mallorca NATO ARW, NATO ASI Series, Springer Verlag, Heidelberg, 3, 157–171, 1992.
Knudsen, K. L.: Foraminiferer i Kvartær stratigrafi: Laboratorie- og fremstillingsteknik samt udvalgte eksempler, Geologisk Tidsskrift, 3, 1–25, 1998.
Knudsen, K. L. and Gibbard, P.: Eemian and Weichselian environmental development in the western Baltic area, NW Europe – introduction, Boreas, 35, 317–319, 2006.
Knudsen, K. L. and Lykke-Andersen, A.-L.: Foraminifera in Late Saalian, Eemian, Early and Middle Weichselian of the Skærumhede I boring, B. Geol. Soc. Denmark, 30, 97–110, 1982.
Knudsen, K. L. and Sejrup, H. P.: Amino acid geochronology of selected interglacial sites in the North Sea area, Boreas, 17, 347–354, 1988.
Knudsen, K. L., Kristensen, P., and Larsen, N. K.: Marine glacial and interglacial stratigraphy in Vendsyssel, northern Denmark: foraminifera and stable isotopes, Boreas, 38, 787–810, 2009.
Knudsen, K. L., Jiang, H., Kristensen, P. Gibbard, P., and Haila, H.: Early Last Interglacial palaeoenvironments in the western Baltic: foraminiferal stable isotopes and diatom-based sea-surface salinity, Boreas, 40, 681–696, 2011.
Knudsen, K. L., Schweizer, M. Seidenkrantz, M.-S., and Pearce, C.: Description and ecology of the new foraminiferal species Elphidium balticum – with a genetic and morphological comparison to Elphidium incertum (Williamson, 1858) and Elphidium asklundi Brotzen, 1943, J. Foramin. Res., Vol. 55, in press, 2024.
Konradi, P., Larsen, B., and Sørensen, A. B.: Marine Eemian in the Danish eastern North Sea, Quatern. Int., 133/134, 21–31, 2005.
Konradi, P. B.: Foraminifera in Eemian deposits at Stensigmose, southern Jutland, Danmarks Geologiske Undersøgelse II. Række, 105, 1–57, 1976.
Kosack, B. and Lange, W.: Das Eem-Vorkommen von Offenbüttel/Schnittlohe und die Ausbreitung des Eem-Meeres zwischen Nord- und Ostsee, Geologisches Jahrbuch, Reihe A, 86, 3–17, 1985.
Kristensen, P. and Knudsen, K. L.: Palaeoenvironments of a complete Eemian sequence at Mommark, southern Denmark: foraminifera, ostracods and stable isotopes, Boreas, 35, 349–366, 2006.
Kristensen, P., Knudsen, K. L., Lykke-Andersen, H., Nørmark, E., Peacock, J. D., and Sinnoth, A.: Interglacial and glacial climate oscillations in a marine shelf sequence from northern Denmark – a multidisciplinary study, Quaternary Sci. Rev., 17, 813–837, 1998.
Kristensen, P., Gibbard, P., Knudsen, K. L., and Ehlers, J.: Last Interglacial stratigraphy at Ristinge Klint, South Denmark, Boreas, 29, 103–116, 2000.
Kubisch, M. and Schönfeld, J.: Eine neue “Cyprinen-Ton”-Scholle bei Stohl (Schleswig-Holstein): Mikrofauna und Grobfraktionsanalyse von Sedimenten der Eem-zeitlichen Ostsee, Meyniana, 37, 89–95, 1985.
Lafrenz, H.-R.: Foraminiferen aus dem marinen Riss-Würm-Interglazial (Eem) in Schleswig-Holstein, Meyniana, 13, 10–46, 1963.
Larsen, E., Sejrup, H. P., Johnsen, S., and Knudsen, K. L.: Do Greenland ice cores reflect NW European Interglacial climate variations?, Quaternary Res., 43, 125–132, 1995.
Larsen, N. K., Krohn, C. F., Kronborg, C., Nielsen, O. B., and Knudsen, K. L.: Lithostratigraphy of the Late Saalian to Middle Weichselian Skærumhede Group in Vendsyssel, northern Denmark, Boreas, 38, 762–786, 2009.
Laskar, J., Robutel, P., Joutel, F., Gastineau, M., Correia, A., and Levrard, B.: A longterm numerical solution for the insolation quantities of the Earth, Astron. Astrophys., 428, 261–285, 2004.
Lauterbach, S., Neumann, F. H., Tjallingii, R., and Brauer, A.: Re-investigation of the Bispingen palaeolake sediment succession (northern Germany) reveals that the Last Interglacial (Eemian) in northern-central Europe lasted at least ∼ 15 000 year, Boreas, 53, 243–261, 2024.
Lutze, G. F.: Zur Foraminiferenfauna der Ostsee, Meyniana, 15, 75–142, 1965.
Lutze, G. F.: Foraminiferen der Kieler Bucht (Westliche Ostsee): 1. “Hausgartengebiet” des Sonderforschungsbereiches 95 der Universität Kiel, Meyniana, 26, 9–22, 1974.
Lykke-Andersen, A.-L. and Knudsen, K. L.: Saalian, Eemian, and Weichselian in the Vendsyssel-Kattegat Region, Denmark, Striae, 34, 135–140, 1991.
Lykke-Andersen, H.: Some main lines of the. Quaternary in the Fennoscandian Border Zone in Kattegat and Vendsyssel, Boreas, 16, 369–371, 1987.
Madsen, V., Nordmann, V., and Hartz, N.: Eem-Zonerne. Studier over Cyprinaleret og andre Eem-aflejringer i Danmark, Nord-Tyskland og Holland, Danmarks Geologiske Undersøgelse II. Række, 17, 1–264, 1908.
Menke, B.: Eem-Interglazial und “Treene-Warmzeit” in Husum/Nordfriesland, Geologisches Jahrbuch, Reihe A, 86, 63–99, 1985.
Menke, B. and Tynni, R.: Das Eeminterglazial und das Weichsel-frühglazial von Rederstall/Dithmarschen und ihre Bedeutung für die mitteleuropäische Jungpleistozän-Gliederung, Geologisches Jahrbuch, Reihe A, 7, 1–120, 1984.
Miller, G. H. and Mangerud, J.: Aminostratigraphy of European marine Interglacial deposits, Quaternary Sci. Rev., 4, 215–278, 1985.
Murray, J.: Ecology and Palaeoecology of benthic foraminifera, Longman, Essex, 397 pp., https://doi.org/10.4324/9781315846101, 1991.
Murray, J.: Ecology and applications of benthic foraminifera, Cambridge University Press, Cambridge, 426 pp., https://doi.org/10.1017/CBO9780511535529, 2006.
Murray, A. S. and Funder, S.: Optically stimulated luminescence dating of a Danish Eemian coastal marine deposit: a test of accuracy, Quaternary Sci. Rev., 22, 1177–1183, 2003.
Müller, H.: Pollenanalytische Untersuchungen und Jahresschichtenzählungen an der eem-zeitlichen Kieselgur von Bispingen/Luhe, Geologisches Jahrbuch, Reihe A, 21, 149–169, 1974.
Ni, S., Quintana Krupinski, N. B., Chonewicz, J., Groeneveld, J., Knudsen, K. L., Seidenkrantz, M.-S., and Filipsson, H. L.: Seasonal climate variations in the Baltic Sea during the Last Interglacial based on foraminiferal geochemistry, Quaternary Sci. Rev., 272, 107–220, https://doi.org/10.1016/j.quascirev.2021.107220, 2021.
Ni, S., Lu, Z., Zhang, Q., Groeneveld, J., Knudsen, K. L., Seidenkrantz, M.-S., and Filipsson, H. L.: Last Interglacial seasonal hydroclimate in the North Sea–Baltic Sea region, Quaternary Sci. Rev., 312, 108–152, https://doi.org/10.1016/j.quascirev.2023.108152, 2023.
Nielsen, J. K., Helema, S., Rodland, D., and Niesen, J. K.: Eemian marine molluks and barnacles from Ristinge Klint, Denmark: hydrodynamics and oxygen deficiency, Neth. J. Geosci., 86, 95–115, 2007.
Nordmann, V.: Molluskfaunaen i Cyprinaleret og Mellem-Europas andre Eem-aflejringer, Danmarks Geologiske Undersøgelse II. Række, 47, 115–264, 1908.
Pearce, E. A., Mazier, F., Normand, S., Fyfe, R., Andrieu, V., Bakels, C., Balwierz, Z., Bińka, K., Boreham, S., Borisova, O. K., Brostrom, A., de Beaulieu, J. L., Gao, C., González-Sampériz, P., Granoszewski, W., Hrynowiecka, A., Kołaczek, P., Kuneš, P., Magri, D., Malkiewicz, M., Mighall, T., Milner, A. M., Möller, P., Nita, M., Noryśkiewicz,.B., Pidek, I. A., Reille, M., Robertsson, A. M., Salonen, J. S., Schläfli, P., Schokker, J., Scussolini, P., Šeirienė, V., Strahl, J., Urban, B., Winter, H., and Svenning, J. C.: Substantial light woodland and open vegetation characterized the temperate forest biome before Homo sapiens, Science Advances, 9, 45, https://doi.org/10.1126/sciadv.adi9135, 2023.
Penney, D. N.: Microfossils (Foraminifera, Ostracoda) from an Eemian (Last Interglacial) tidal flatsequence in South-West Denmark, Quatern. Int., 3–4, 85–91, 1989.
Roebroeks, W., Gibbard, P. L., and Scherjon, F.: Taphonomy matters: Comment on “Substantial light woodland and open vegetation characterized the temperate forest biome before Homo sapiens” by Pearce et al. 2023, Science Advances [preprint], https://doi.org/10.31219/osf.io/ub572, 2024.
Rosenfeld, A.: Die rezenten Ostracoden-Arten in der Ostsee, Meyniana, 29, 11–49, 1977.
Sánchez Goñi, M. F., Crucifix, M., Peyron, O., Santos, L., Duprat, J., Malaize, B., Turon, J.-L., and Peypouquet, J.-P.: Increasing vegetation and climate gradient in Western Europe over the Last Glacial Inception (122–110 ka): data-model comparison, Earth Planet. Sc. Lett., 231, 111–130, 2005.
Seidenkrantz, M.-S.: Benthic foraminiferal and stable isotope evidence for a “Younger Dryas-style” cold spell at the Saalian-Eemian transition, Denmark, Palaeogeogr. Palaeocl., 102, 103–120, 1993a.
Seidenkrantz, M.-S.: Foraminifera from the Quaternary sequence in the Anholt boring, Denmark, Boreas, 22, 283–290, 1993b.
Seidenkrantz, M.-S., Knudsen, K. L., and Kristensen, P.: Marine late Saalian to Eemian environments and climatic variability in the Danish shelf area, Neth. J. Geosci., 79, 335–343, 2000.
Shackleton, N. J.: Oxygen isotopes, ice volume and sea level, Quaternary Sci. Rev., 6, 183–190, 1987.
Shackleton, N. J.: The 100,000-year ice age cycle identified and found to lag temperature, carbon dioxide, and orbital eccentricity, Science, 289, 1897–1902, 2000.
Shackleton, N. J., Sánchez Goñi, M. F., Pailler, D., and Lancelot, Y.: Marine Isotope Substage 5e and the Eemian Interglacial, Global Planet. Change, 36, 151–155, 2003.
Thiel, C., Kenzler, M., Stephan, H.-J., Frechen, M., Urban, B., and Sierralta, M.: Chronological and sedimentological investigations of the Late Pleistocene succession in Osterbylund (Schleswig-Holstein, Germany), E&G Quaternary Sci. J., 72, 57–72, https://doi.org/10.5194/egqsj-72-57-2023, 2023.
Turner, C.: Problems of the duration of the Eemian interglacial in Europe north of the Alps, Quaternary Res., 58, 45–48, 2002.
Turner, C. and West, R. G.: The subdivision and zonation of interglacial periods, E&G Quaternary Sci. J., 19, 93–101, https://doi.org/10.3285/eg.19.1.06, 1968.
West, R. G.: Pleistocene geology and biology, with especial reference to the British Isles, 2nd edn., Longman, London, 377 pp., ISBN-13. 978-0582446205, 1977.
West, R. G.: Pleistocene Forest History in East Anglia, New Phytol., 85, 571–622, 1980.
Winn, K., Glos, R., Averdieck, F.-R., and Erlenkeuser, H.: On the age of the marine Eem in northwestern Germany, Geologos, 5, 41–56, 2000.
Zagwijn, W. H.: Sea-level changes in the Netherlands during the Eemian, Neth. J. Geosci., 62, 437–450, 1983.
Zagwijn, W. H.: An analysis of Eemian climate in Western and Central Europe, Quaternary Sci. Rev., 15, 451–469, 1996.